

33. Взаимодействие бактериофагов с бактериальной клеткой. Вирулентные и умеренные бактериофаги.
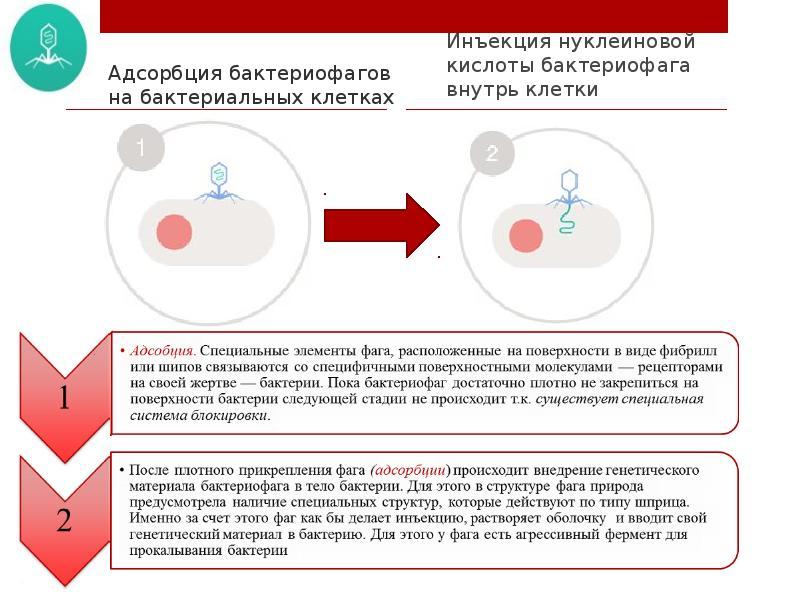


Процесс взаимодействия вирулентного фага с бактерией протекает в виде нескольких стадий и весьма схож с процессом взаимодействия вирусов человека и животных с клеткой хозяина:
1. Первая стадия - адсорбция фага на рецепторные участки клеточной стенки бактерий. К ним фаг прикрепляется концевыми нитями отростков. Адсорбция фага на чувствительной клетке. Происходит при наличии комплементарных рецепторов в клеточной стенке бактерий или на концах нитей фагового отростка
2. Проникновение - на этой стадии ДНК фага через отросток проникает в клетку. При этом слои клеточной стенки разрушаются под действием фагового лизоцима. Проникновение ДНК фага в бактериальную клетку. С помощью лизоцима осуществляется гидролиз участка клеточной стенки, чехол отростка сокращается и внутренний стержень прокалывает оболочку клетки. ДНК по каналу стержня проникает внутрь
3. Третья стадия - на этой стадии начинается биосинтез фаговой информационной РНК, белков капсида, которые участвуют в биосинтезе фаговой ДНК. Латентный период продолжается в пределах 15 минут. Внутриклеточное развитие фага. ДНК бактериофага направляет клеточные системы на биосинтез компонентов, необходимых для репродукции фагов. Сначала идет синтез «ранних белков» – ферментов, осуществляющих репликацию ДНК, а затем «поздних белков» – белков головки, отростка и т.д.
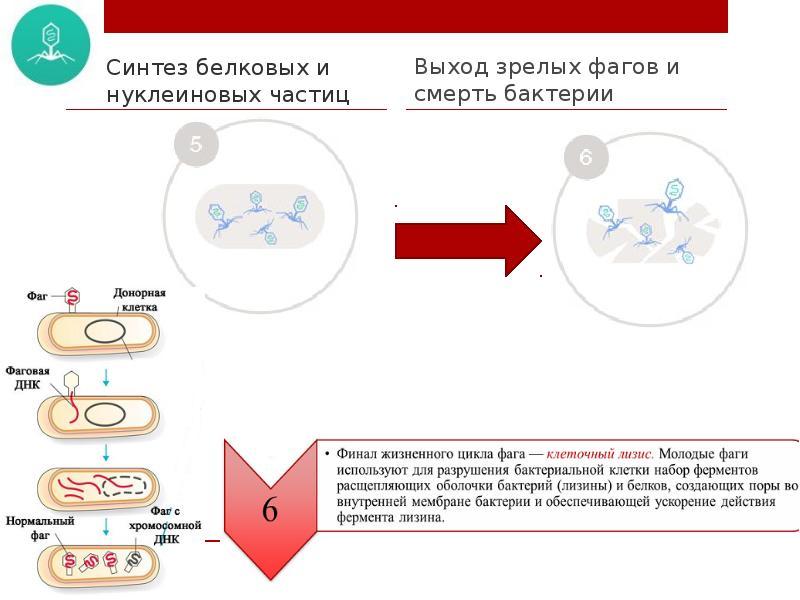
4. Четвертая стадия - морфогенез фага, т.е. пустотелые фаговые капсиды заполняются нуклеиновой кислотой и формируются зрелые вирионы (частицы фага). Морфогенез фага. Созревание фага - разобщенный процесс. Отдельно формируются головки фага: вокруг ДНК строится капсид. Независимо образуется отросток: формируется базальная пластинка, к ней прикрепляется внутренний стержень и одевается чехлом. Отдельно синтезируются нити отростка. Затем составные части фага объединяются, образуя вирионы
5. Пятая стадия - выход фаговых частиц из клетки. Это происходит благодаря лизису зараженной бактерии фаговым лизоцимом, накапливающимся в процессе репродукции фага. Лизис бактериальной клетки и выход фага. Фаговый лизоцим гидролизует клеточную стенку и осуществляет лизис клетки. Бактериофаги выходят в окружающую среду
Количество зрелых фаговых частиц (вирионов) колеблется от единиц до нескольких тысяч. Затем фаги вновь внедряются в еще незараженные клетки и процесс повторяется.
растворение (лизис).
Взаимодействие фага с бактериальной клеткой. По механизму взаимодействия различают вирулентные и умеренные фаги.
Различают два типа взаимодействия фага с клеткой:
1) литический (продуктивная вирусная инфекция). Это тип взаимодействия, при котором происходит репродукция вируса в бактериальной клетке. Она при этом погибает.
2) лизогенный. Это умеренные фаги. При проникновении нуклеиновой кислоты в клетку идет интеграция ее в геном клетки, наблюдается длительное сожительство фага с клеткой без ее гибели. При изменении внешних условий могут происходить выход фага из интегрированной формы и развитие продуктивной вирусной инфекции.
Вирулентные фаги, проникнув в бактериальную клетку, автономно репродуцируются в ней и вызывают лизис бактерий. Процесс взаимодействия вирулентного фага с бактерией протекает в виде нескольких стадий и весьма схож с процессом взаимодействия вирусов человека и животных с клеткой хозяина. Однако для фагов, имеющих хвостовой отросток с сокращающимся чехлом, он имеет особенности. Эти фаги адсорбируются на поверхности бактериальной клетки с помощью фибрилл хвостового отростка. В результате активации фагового фермента АТФазы происходит сокращение чехла хвостового отростка и внедрение стержня в клетку. В процессе «прокалывания» клеточной стенки бактерии принимает участие фермент лизоцим, находящийся на конце хвостового отростка. Вслед за этим ДНК фага, содержащаяся в головке, проходит через полость хвостового стержня и активно впрыскивается в цитоплазму клетки. Остальные структурные элементы фага (капсид и отросток) остаются вне клетки.
После биосинтеза фаговых компонентов и их самосборки в бактериальной клетке накапливается до 200 новых фаговых частиц. Под действием фагового лизоцима и внутриклеточного осмотического давления происходит разрушение клеточной стенки, выход фагового потомства в окружающую среду и лизис бактерии. Один литический цикл (от момента адсорбции фагов до их выхода из клетки) продолжается 30—40 мин. Процесс бактериофагии проходит несколько циклов, пока не будут лизированы все чувствительные к данному фагу бактерии.
Взаимодействие фагов с бактериальной клеткой характеризуется определенной степенью специфичности. По специфичности действия различают поливалентные фаги, способные взаимодействовать с родственными видами бактерий, моновалентные фаги, взаимодействующие с бактериями определенного вида, и типовые фаги, взаимодействующие с отдельными вариантами (типами) данного вида бактерий.
Умеренные фаги лизируют не все клетки в популяции, с частью из них они вступают в симбиоз, в результате чего ДНК фага встраивается в хромосому бактерии. В таком случае геномом фага называют профаг. Профаг, ставший частью хромосомы клетки, при ее размножении реплицируется синхронно с геном бактерии, не вызывая ее лизиса, и передается по наследству от клетки к клетке неограниченному числу потомков.
Умеренные фаги, в отличие от вирулентных, не всегда вызывают гибель бактериальных клеток и при взаимодействии с ней переходят в неинфекционную форму фага, называемую профагом. Умеренные фаги, в отличие от вирулентных, не всегда вызывают гибель бактериальных клеток и при взаимодействии с ней переходят в неинфекционную форму фага, называемую профагом. Профаг — геном фага, ассоциированный с бактериальной хромосомой. Профаг, ставший частью хромосомы клетки, при ее размножении реплицируется синхронно с геномом бактерии, не вызывая ее лизиса, и передается по наследству от клетки к клетке в неограниченном числе поколений.
Биологическое явление симбиоза микробной клетки с умеренным фагом (профагом) называется лизогенией, а культура бактерий, содержащая профаг, получила название лизогенной. Это название отражает способность профага самопроизвольно или под действием ряда физических и химических факторов исключаться из хромосомы клетки и переходить в цитоплазму, т. е. вести себя как вирулентный фаг, лизирующий бактерии.
Лизогенные культуры по своим основным свойствам не отличаются от исходных, но они невосприимчивы к повторному заражению гомологичным или близкородственным фагом и, кроме того, приобретают дополнительные свойства, которые находятся под контролем генов профага. Изменение свойств микроорганизмов под влиянием профага получило название фаговой конверсии. Последняя имеет место у многих видов микроорганизмов и касается различных их свойств: культуральных, биохимических, токсигенных, антигенных, чувствительности к антибиотикам и др. Кроме того, переходя из интегрированного состояния в вирулентную форму, умеренный фаг может захватить часть хромосомы клетки и при лизисе последней переносит эту часть хромосомы в другую клетку. Если микробная клетка станет лизогенной, она приобретает новые свойства. Таким образом, умеренные фаги являются мощным фактором изменчивости микроорганизмов.
Умеренный бактериофаг сохраняет режим профага, следует лизогенному пути. Вирулентные представители развиваются в соответствие с литической моделью, в которой имеется ряд последовательных процессов: Направление синтеза нуклеиновых кислот задается ферментами фага, который влияет на аппарат, отвечающий за белковый синтез. Паразит начинает инактивацию РНК и ДНК, принадлежащих хозяину, а дальнейшее ферментативное воздействие вовсе приводит к ее расщеплению. На следующей части процесса происходит «подчинение» клеточного аппарата по белковому синтезу. Фаговая н. к. подвергается репликации и обуславливает направление синтеза новых белковых оболочек. Процесс образования лизоцима находится в подчинении фаговой РНК. Клеточный лизис: разрыв клетки, обусловленный деятельностью лизоцима. Происходит высвобождение огромного числа новых фагов, которые будут инфицировать бактериальные организмы дальше. Поливалентные бактериофаги взаимодействуют с родственными видамибактерий; Моновалентные – с бактериями одного типа; Типовые – с отдельными вариантами бактерий данного вида
ФАГОВАЯ/лизогенная КОНВЕРСИЯ — один из видов изменчивости бактерий, обусловленный геномом бактериофага, внедрившимся в полинуклеотидную последовательность ДНК бактериальной клетки. Приобретение клеткой нового фенотипического признака в результате фаговой конверсии не связано с нарушением имеющейся в ней генетической информации (см. Бактерии, генетика), а является следствием добавления к геному бактерий генов бактериофага. (например, появление способности к образованию экзотоксина у возбудителей ботулизма, дифтерии, скарлатины, холеры).