

Биохимия возникновения и проведения нервного импульса
Клетки сообщаются друг с другом путём передачи сигналов, которые регистрируются в виде электрических процессов. Характер изменения мембранного потенциала постсинаптической клетки зависит от типа канала.
-
Изменение мембранного потенциала от -60 до +30 мВ за счёт открывания Nа+-каналов ведет к возникновению постсинаптического потенциала действия.
-
Изменение мембранного потенциала с -60 мВ до -90 мВ за счёт открывания Cl–-каналов ингибирует потенциал действия (гиперполяризация), в результате возбуждение не передается (тормозной синапс).
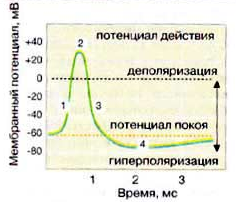
Рис. 6. Мембранный потенциал постсинаптической клетки
В состоянии покоя внутренняя сторона мембраны нейрона заряжена отрицательно. Это потенциал покоя, величина его достигает 60-75 мВ. Он обеспечивается в основном работой натриевого насоса (Na+,К+-АТФазы). За счет энергии гидролиза каждой молекулы АТФ три иона Na+ выкачиваются из клетки во внеклеточное пространство, а два иона К+ перемещаются в противоположном направлении.
Разность потенциалов возникает в основном за счет следующих явлений. Несмотря на то что мембрана достаточно проницаема для ионов К+, их концентрация внутри аксона в 20-50 раз выше, чем во внешней среде. Главные внутриклеточные анионы (белки и нуклеиновые кислоты) не могут выходить наружу, а ионы С1–, которых много во внешней среде, проходят через мембрану очень медленно. Проницаемость для ионов Na+ составляет лишь 1/20 по сравнению с проницаемостью для ионов К+. При таких условиях трансмембранный потенциал определяется распределением ионов К+. Этот потенциал существует благодаря тому, что ионы К+ стремятся покинуть клетку, чтобы уравнять внешнюю и внутреннюю концентрации. Однако в клетке при этом остается избыток анионов, что создает отрицательный электрический заряд, ограничивающий дальнейшее выравнивание концентраций ионов К+. Ионы С1– ведут себя наоборот: они должны остаться снаружи, чтобы сбалансировать электрический заряд плохо проникающего Na+, но в то же время стремятся проникнуть в клетку по градиенту концентрации.
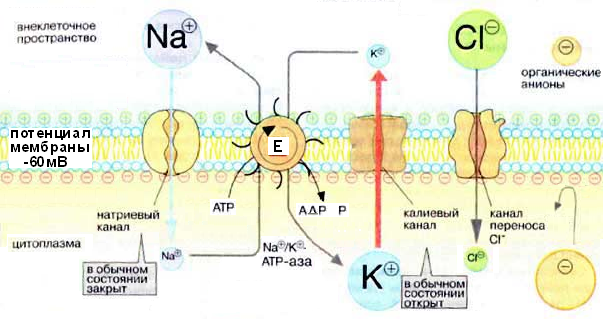
Рис. 6. Механизмы поддержания мембранного потенциала покоя в постсинаптической клетке
При возбуждении, вызванном тем или иным агентом (например, электрическим стимулом), избирательно увеличивается проницаемость мембраны нервной клетки (аксона) для ионов Na+. Некоторое количество ионов Na+ устремляется внутрь клетки. В результате возникает «овершут», т.е. величина потенциала может изменяться от –75мВ до +30 мВ на внутренней поверхности мембраны. Этот положительный заряд препятствует дальнейшему входу Na+, проводимость для Na+ падает, a Na+-нacoc восстанавливает исходное состояние. Эта последовательность процессов называется потенциалом действия и завершается примерно в течение 1 мс. Многочисленные Nа+-каналы миелинизированных волокон сосредоточены в немиелинизированных перехватах Ранвье. Под миелиновой оболочкой относительно длинных межперехватных участков очень мало натриевых каналов. Деполяризация одного из перехватов вызывает градиент потенциала между перехватами, благодаря которому через аксоплазму быстро протекает ток к соседнему перехвату, вызывая снижение потенциала до порогового уровня. В итоге получается высокая скорость проведения импульса по миелинизированному волокну.
За один импульс через мембрану проходит небольшое количество ионов Na+ и К+, и концентрационные градиенты обоих ионов сохраняются: в клетке выше уровень К+, а вне клетки выше уровень Na+. Поэтому по мере получения клеткой новых импульсов процесс локального обращения знака мембранного потенциала может повторяться многократно.
Распространение потенциала действия по поверхности нервной клетки основано на том, что локальное обращение мембранного потенциала стимулирует открывание соседних потенциал-управляемых ионных каналов, в результате чего возбуждение распространяется в виде деполяризационной волны на всю клетку.