3. Хромомерная организация хромосом
В конце 19-го века ряд исследователей (E.G. Balbiani, W. Pfitzner, W. Flemming) (см. Вильсон, 1936) нашли, что хромосомная нить на стадии профазы митоза содержит ряд небольших сильно окрашивающихся телец или хромомеров, различающихся по размерам и форме. В профазе мейоза рисунок хромомеров в гомологичных хромосомах полностью симметричен.
"Каждое хроматиновое зернышко одной нити имеет двойника в другой и нет хотя бы малейшей особенности одной нити, которая не повторялась быв точности у ее спутника" (Вильсон, 1936). Различаясь между собой по величине, форме, содержанию ДНК и положению в хромосоме, хромомеры обладают отчетливо выраженной индивидуальностью и придают хромосомным нитям и отдельным их участкам постоянный и определенный рисунок, строго фиксированный наследственно (Прокофьева-Бельговская, Богданов, 1963).
Только в профазе митоза и мейоза (лептотена и пахитена) хромосомы имеют хромомерную организацию у всех эукариот, на других стадиях клеточного цикла хромомеры встречаются довольно редко. Идеи о том, что хромомеры (или гранулярность хромосомной нити) существуют и в интерфазных ядрах, в которых индивидуальные хромосомы не опознаются, развивал еще В. Флеминг в 1882 г.
Наиболее выраженные хромомеры встречаются в политенных хромосомах и хромосомах типа "ламповых щеток". От хромомеров отходят боковые петли, в результате чего хромосома выглядит как ершик для чистки керосиновых ламп, поэтому их и называют "хромосомами типа ламповых щеток" - lampbrash chromosomes.
У. Дюрие в 1941 году описал хромомеры в хромосомах типа "ламповых щеток", которые находят в первичных ооцитах (профаза, по-видимому, диплотена) многих позвоночных и некоторых беспозвоночных
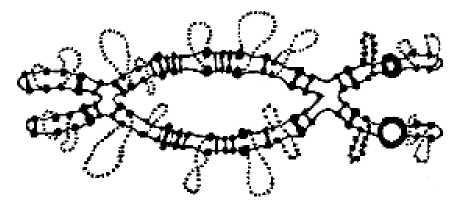
Два свойства являются общими для хромомеров различных типов: во-первых, все они представляют собой отрезки компактизованной ДНК, во-вторых, число и рисунок хромомеров у данного организма постоянны на данной стадии клеточного цикла.
Таким образом, завершая общее рассмотрение понятия хромомера, можно подчеркнуть, что это фрагменты хромосом, способные к локальной компактизации. Длина фрагмента ДНК, входящего в состав хромомера, различна на разных этапах процесса компактизации хромосом в ходе клеточного цикла. Поэтому есть все основания считать, что хромомер не является постоянной структурной единицей организации генома, и для каждого этапа онтогенеза существуют свои наборы хромомеров. Поэтому все известные типы хромомеров можно разделить, по крайней мере, на четыре группы: а) лептотенные, б) пахитенные, в) хромосом типа "ламповых щеток", г) хромомеры интерфазных политенных хромосом.
4. Митотические хромосомы
Еще в 1882 году Сграсбургером было обнаружено у одного из исследованных им растений, а именно Funkia sieboldiana, что хромосомы одной и той же ядерной пластинки весьма резко отличаются по своей величине. Аналогичные отношения были констатированы впоследствии для целого ряда растений и животных. Несколько позже, Мюллер (Cl. Muller, 1912) посвятил целое исследование, специально посвященное различиям в размерах хромосом. Указанные различия являлись не простым варьированием, а характеризовали собой определенные типы более крупных и мелких хромосом, точно повторявшиеся в различных ядерных пластинках одного и того же вида. Каждый тип был представлен в соматических клетках парой одинаковых элементов, очевидно отцовского и материнского происхождения. Такого рода данные представляли - рядом с постоянством числа хромосом - наглядную иллюстрацию индивидуальности хромосом, понимаемой в смысле определенных, характерных для каждой пары особенностей.
Точным установлением факта, что хромосомам присущи, помимо их абсолютной и относительной величины, еще и особые постоянные и характерные различия в построении их тела, наука обязана трудам Сергея Гавриловича Навашина(1910-1914).
Уже в ранних работах Навашин выделяет три типа хромосом: a) U-образные, почти равноплечие, б) U-образные, явственно неравноплечие, в) крючковидные, один членик которых настолько короток, что может даже ускользнуть от наблюдения (Навашин, 1911).
В 1912 году на заседании физико-математического отдела Академии наук состоялось знаменитое сообщение С.Г. Навашина, где он установил у неоднократно до того подвергавшегося исследованию обьекта Galtonia candicans наличие особых мельчайших, но вполне постоянных придатков, присоединенных при помощи "ниточки" к двум "средним" хромосомам. Придатки эти были названы С.Г. Навашиным "спутниками" (satelles - лат). Тельца эти при делении ядра расщепляются вместе с остальным телом хромосомы. Таким образом, впервые была показана возможность идентификации хромосом по особенностям их строения.
В 1914 году С.Г. Навашин установил, что в участке прикрепления нитей веретена образуется перетяжка материала хромосомы и эта перетяжка расположена в характерных местах в трех ранее установленных типах хромосом.
Из-за того, что данные Навашина были опубликованы на русском языке, к тому же в специальных изданиях, а также из-за последовавших вскоре политических пертурбаций, его работа осталась совершенно неизвестной за границей. Факты, установленные Навашиным, постепенно открывались иностранными учеными вторично, например, много позже Ньютоном и Тейлором. (Newton, 1924; Taylor, 1924). Оба они, как мы видим, с большим отставанием открыли спутники и перетяжки хромосом в месте прикрепления нитей веретена. (строение хромосомы)
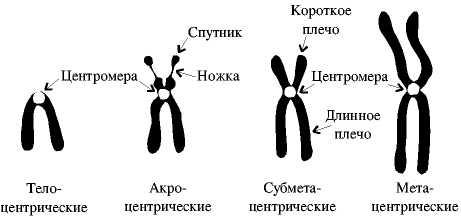
Фактически в соответствии с классификацией Навашина, выделяют 4 типа хромосом в зависимости от положения центромеры и определяемой этим положением относительной длины плеч, т.е. частей хромосомы по обе стороны от центромеры.
По мнению многих ученых любая хромосома имеет два плеча, т.е. телоцентрической хромосомы в природе не существует. У телоцентрических хромосом во всех случаях обнаружено наличие второго, пусть очень короткого плеча.