

3. Трансляция
Трансляция(перевод) – процесс воплощения генетической информации мРНК в структуру полипептида. Это второй этап белкового синтеза, осуществляемый последовательной поликонденсацией отдельных аминокислотных остатков, начиная с аминоконца полипептидной цепи к карбоксильному концу.
Зрелая матричная РНК выходит в цитоплазму, где осуществляется процесс транскрипции – декадирование мРНК в аминокислотную последовательность белка. Процесс декадирования осуществляется в направлении от 5`→3` и происходит в рибосомах. Комплекс мРНК и рибосом называется полисомой. Подобно транскрипции механизм трансляции состоит из трех этапов: инициации, элонгации и терминации.
Трансляция начинается со стартового кодона АУГ, который при локализации в смысловой части структурного гена кодирует аминокислоту метионин. Каждую аминокислоту доставляет к полисоме транспортная РНК (тРНК), специфичная к данной аминокислоте. тРНК выполняет роль посредника между кодоном мРНК и аминокислотой. Молекулы тРНК узнают в цитоплазме соответствующий триплет (кодон в мРНК) по принципу спаривания комплементарных азотистых оснований. тРНК, которая подходит к малой субчастице, образует связь кодон-антикодон, при этом одновременно передает свою аминокислоту в аминоацильный участок (А-участок) большой субъединице. К кодону АУГ «подходит» антикодон только той тРНК, которая переносит метионин. Поэтому прежде всего к рибосоме доставляется метионин. Затем кодон АУГ переходит на пептидильный участок большой субъединицы (Р-участок). В результате этих процессов образуется транслирующая рибосома – инициирующий комплекс.
Элонгация– это последовательное включение аминокислотных остатков в состав растущей полипептидной цепи. Каждый акт элонгации состоит из трех этапов:
- узнавание кодона, которое заключается в связывании антикодона с очередной молекулой тРНК, несущей аминокислоту, с кодоном свободного А-участка на рибосоме;
- образование пептидной связи, которое происходит лишь тогда, когда оба участка А и Р заняты молекулами тРНК. Часть большой субъединицы рибосомы – фермент пептидилтрансферазу, катализирующий образование пептидной связи;
- транслокация, где тРНК участка Р, не связанная с пептидом, покидает рибосому. Затем молекула тРНК с полипептидом переходят из А на Р-участок и, наконец, рибосома перемещается вдоль РНК на один кодон.
Терминация(окончание синтеза) происходит по команде кодонов УАА, УАГ, УГА. В природе не существует таких молекул тРНК, антикодоны которых соответствовали бы этим кодонам.
Каждая мРНК транслируется, как правило, несколько раз, после чего разрушается. Среднее время жизни молекулы мРНК около 2 мин. Разрушая старые и образуя новые мРНК, клетка может довольно строго регулировать как тип продуцируемых белков, так и их количество. Это регуляция синтеза белка на уровне транскрипции. У эукариот возможна регуляция и на уровне трансляции.
Синтез белка – один из существеннейших показателей жизни.
4. Передача информации в клетке
Современные представления о роли ДНК в передаче наследственной информации лучше всего отражает "Центральная догма молекулярной биологии", сформулированная Ф. Криком в 1970 году.
Автор предложил разделить все виды переноса биологической информации в клетке на три группы:
1. Процессы, существование которых уже показано: ДНК → ДНК, ДНК → РНК, РНК → белок, РНК → РНК.
2. Процессы, которые не были экспериментально выявлены и с теоретической точки зрения не казались строго необходимыми: РНК → ДНК, ДНК → белок.
3. Невозможные переносы: белок → белок, белок → РНК, белок → ДНК. Таким образом, информация во всех случаях в клетке переносится однонаправленно по цепи: ДНК →РНК → белок. Белок не может служит матрицей для синтеза ДНК или РНК, поскольку у молекул белка нет свойства комплементарности отдельных частей молекулы, что бы позволяло использовать её как матрицу.
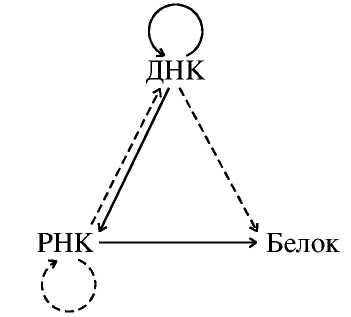
"Центральная догма молекулярной биологии". Сплошные стрелки показывают обычный путь переноса генетической информации, пунктирной - более редкие пути, также существующие в природе
Синтез одной молекулы белка, состоящего из 150 аминокислот, идет примерно за 1,5 минуты, т. е. со скоростью 2 аминокислоты в секунду. Он зависит от многих факторов. Например, состояние рибосомы может оказать влияние на считывание информации. Рибосома «читает с ошибками», если на нее воздействовать какими-либо внешними факторами, к примеру облучением, химическими веществами, способными изменять структуру и функцию рибосомы.
Однако если переносы типа ДНК→ДНК, ДНК→РНК, РНК→РНК и РНК→белок имели экспериментальные прямые или непрямые доказательства, то в пользу других переносов доводов и теоретических обоснований в то время не было.
Изучение механизмов взаимодействия с клеткой опухолеродных вирусов натолкнуло на мысль о возможности существования иных типов связей. В 1969-1971 гг. Р. Дульбекоэкспериментально доказал, что ДНК опухолеродного вируса прочно связывается с ДНК клетки, находя в ее хромосомах тайное убежище. Но опухолеродные вирусы делятся на две большие группы: ДНК-содержащие и РНК-содержащие. Включение вирусной ДНК в ДНК клетки и их интеграцию представить легко. Но как применить эту гипотезу к РНК-содержащим вирусам?
В 60-х годах Г. Теминвысказал предположение, согласно которому «жизненный цикл» РНК-содержащих опухолеродных вирусов должен включать стадию образования ДНК-продукта — провируса. Это явно противоречило центральной догме молекулярной биологии, гласившей, что генетическая информация передается тольков одном направлении: ДНК→РНК→белок. Если допустить, что существует и путь РНК→ДНК, то в клетке должен быть и специальный фермент, участвующий в синтезе такого рода.В 1970 г. Г. Темин и С. Мизутаниобнаружилив составе вируса саркомы Рауса (РНК-содержащий вирус)фермент, способный синтезировать ДНК на матрице РНК.Этот фермент назвали обратной транскриптазой или РНК-зависимой ДНК-полимеразой (ревертазой, по В. А. Энгельгардту). Одновременно Д. Балтимор обнаружил фермент, синтезирующий ДНК, у вируса миелобластоза птиц. Ревертаза в настоящее время найдена во всех без исключения РНК-содержащих опухолеродных вирусах.В 1975 г. Р. Дульбеко, Г. Темину и Д. Балтимору была присуждена Нобелевская премия за открытие процесса передачи наследственной информации, который получил название обратной транскрипции. Формула центральной догмы молекулярной биологии дополнилась:
ДНК ← → РНК → белок
Процесс обратной транскрипции состоит из двух этапов. Сначала на РНК-матрице синтезируется нить ДНК, т. е. образуется промежуточный продукт реакции, состоящий из гибридных молекул, одна нить которых — вирусная РНК, другая — комплементарная ей синтезированная нить ДНК. На ДНК-вой нити гибридной молекулы синтезируется вторая нить ДНК и получается конечный продукт реакции — двухцепочечная спиральная молекула ДНК, содержащая генетическую информацию, полностью переписанную с вирусной РНК.Оба этапа осуществляются, по-видимому, одним и тем же вирусным ферментом, точнее, его активным центром, т. е. ревертаза обладает и РНК-зависимой и ДНК-зависимой ДНК-полимеразной активностью и ведет всю реакцию от начала до конца.
Открытие ревертазы натолкнуло на мысль, что путем выявления ее в клетках можно осуществлять раннюю и быструю диагностику злокачественных опухолей и лейкозов. Однако вскоре обнаружилось, что ревертазы свойственны и тканям здоровых, не зараженных вирусами организмов. Особенно много их оказалось в эмбриональных клетках. Тем не менее существуют количественные различия ревертаз в опухолевых и нормальных клетках, за исключением эмбриональных. Установлены также различия в активности и физико-химических свойствах ревертаз онкогенных вирусов и нормальных клеток. Несомненно, обратная транскрипция нужна для злокачественной трансформации.
Наличие ревертазы во всех нормальных клетках свидетельствует о возможности передачи информации от РНК к ДНК. Но с какой целью? Отмечено, что на определенной стадии эмбриогенеза в клетках амфибий резко возрастает число генов, кодирующих рибосомальную РНК. Вместо двух копий (2 гомологичные хромосомы) в клетках обнаруживается по несколько сотен копий каждого гена, которые определенный период эмбриогенеза функционируют изолированно от хромосомы, а затем разрушаются. В 1971 г. Тартоф открыл такое же явление и у дрозофилы. Оно былоназвано амплификацией генов. Механизм амплификации не известен. Однако установлено, что в условиях повышенного требования синтеза белка в клетке происходит размножение генов рибосомальной РНК методом обратной транскрипции. Это обеспечивает синтез РНК не на хромосомной матрице, а на матрице генов, образующихся в цитоплазме. Очевидно, амплификация генов играет существенную роль в регуляции феногенетических процессов и происходит всегда, когда требуется увеличить количество белка.
Литература
Айала, Ф. Современная генетика / Ф. Айала, Дж. Кайгер. – М.: Мир, 1987. – Т.1. – 295 с; Т.2. – 368 с; Т.3.
Алиханян, С. И. Общая генетика / С. И. Алиханян, А. П. Акифьев, Л. С. Чернин. – М.: Высш. шк., 1985.
Бокуть, С. Б. Молекулярная биология: молекулярные механизмы хранениия, воспроизведения и реализации генетической информации / С. Б. Бокуть, Н. В. Герасимович, А. А. Милютин. – Мн.:Высш. шк., 2005.
Дубинин, Н. П. Общая генетика / Н. П. Дубинин. – М.: Наука, 1986.
Жимулев, И. Ф. Общая и молекулярная генетика / И. Ф. Жимулев. – Новосибирск: Изд-во Новосибирского ун-та, 2002.
Жученко, А. А. Генетика / А. А Жученко, Ю. Л. Гужов, В. А. Пухальский. – М.: Колос, 2004.