

Билатеральный гинандроморф y Drosophila melanogastei
признакам между самцами и самками, т.е. к появлению интерсексуальности. Интерсексы обнаружены и у дрозофилы.
От гинандроморфов интерсексы отличаются тем, что у них отсутствуют различно детерминированные по полу сектора.
У интерсексов до определенного момента развития сохраняется генетически детерминированный пол, но затем развитие продолжается в направлении противоположного пола. В результате интерсексы отличаются от нормальных особей тем, что у них первичные и вторичные половые признаки носят промежуточный характер, образуя непрерывный ряд переходов от нормального самца к нормальной самке.
Наряду с разнополостью у многих растений и у низших животных мужской и женский пол совмещается в одном организме, который таким образом является гермофродитом.
4. Наследование признаков сцепленных с полом.
Наследование признаков, гены которых находятся в X и У хромосомах, называется наследованием, сцепленным с полом.
Т.Г. Морган и его сотрудники провели два типа скрещиваний дрозофил: в одном самки были нормальными по цвету глаз (w+), а самцы имели белые глаза (w), в другом белоглазых самок (w) скрещивали с нормальными самцами
(w ). Такие скрещивания называют реципрокными, т.е. проведенные в обоих направлениях. В скрещивании нормальных самок с белоглазыми самцами
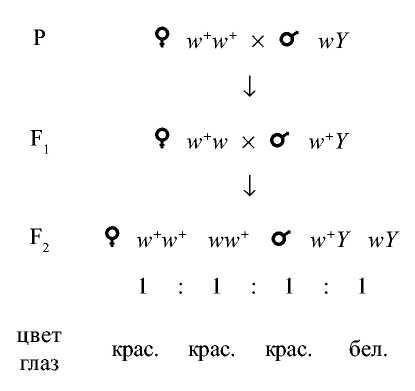
все самцы и самки первого поколения были красноглазыми (w+w и w+Y). Во втором поколении все самки были красноглазыми, а самцы - красноглазыми и белоглазыми, в соотношении 1:1.
Расщепление 3:1 получается, но своеобразное, самки все одного фенотипа, а самцы - двух.
В случае реципрокного скрещивания, когда самка, гомозиготная по гену w (белые глаза), скрещивается с красноглазым самцом, расщепление наблюдается в первом же поколении в отношении белоглазых и красноглазых 1:1.
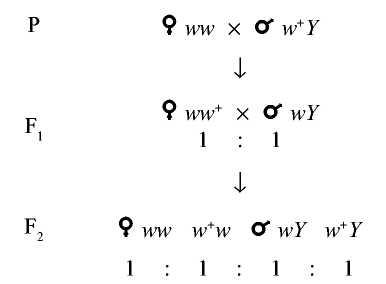
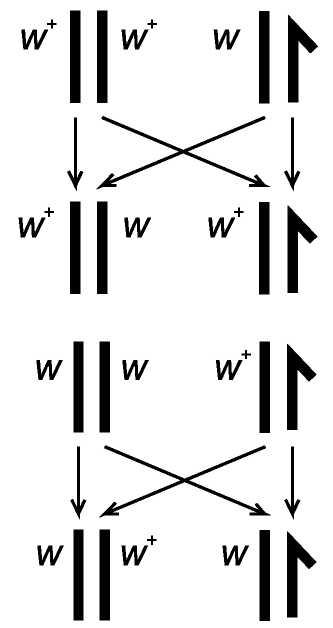
Схема передачи X-хромосомы от самки w+/w+ и w/w самцу и наследование крисс-кросс
При этом белоглазыми оказываются только самцы, а все самки - красноглазые. В F2 и самки, и самцы представлены и белоглазыми, и красноглазыми особями в равных соотношениях.
Картина наследования, когда в F1 признаки родителей передаются противоположному полу, называется крисс-кросс (criss-cross - крест-накрест).
Эти расщепления полностью коррелируют с поведением половых хромосом. Именно в этом эксперименте была впервые показана генетическим образом роль хромосом в наследственности.
5.Сцепление генов и кроссинговер. Генетические доказательства перекреста хромосом
Как и в других законах наследственности, в законе о сцеплении генов сразу же обнаружили исключения. Морган в 1911 году нашел, что в гомологичной паре хромосом регулярно происходит обмен генами.
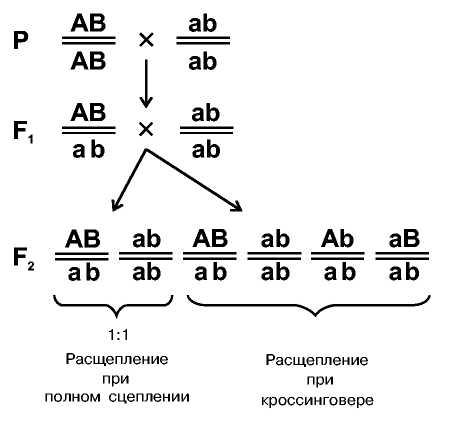
В скрещивании организмов, различающихся по паре признаков, в F1 получаются дигетерозиготы АВ/аb.
В скрещивании потомков F1 с родительской формой ab/ab в случае полного сцепления получалось бы расщепление АВ/ab и ab/ab в соотношении 1:1. Однако, всегда появляются новые сочетания признаков, например, Ab/ab и aB/ab. Значит, во время гаметогенеза образовались новые сорта гамет за счет перекреста хромосом и обмена их фрагментами.
Дополнение
Отмечая заслуги Т.Х. Моргана в формулировании хромосомной теории наследственности, в 1933 году ему была присуждена Нобелевская премия.
Т.Х. Морган с сотрудниками скрещивал линии дрозофил, содержащие гены a – черное тело, b - зачаточные крылья).
Далее, ставились реципрокные скрещивания: в одном дигетерозиготой была самка, а дигомозиготой - самец, в другом скрещивании - наоборот.
Если дигетерозиготой был самец, в потомстве 1 часть имеет фенотип Ab, другая часть - aB. Эти классы расщепляются в соотношении 1:1.
В реципрокном скрещивании получено четыре класса потомков, два из которых имеют сцепленные гены, в том порядке, в каком они наблюдались у родителей, а два других класса возникли в результате нарушения сцепления - это кроссоверы:
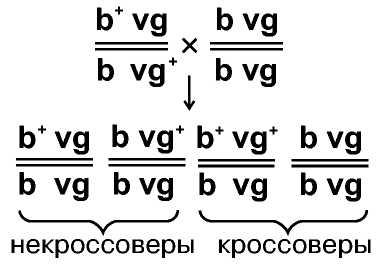
Эти результаты неопровержимо показывают, что в ходе гаметогенеза произошел обмен фрагментами хромосом.
В каждом из классов число мух было в определенных числовых сотношениях: Ab/ab и aB/ab составляли по 41,5%, т.е. некроссоверов было 83%. Два кроссоверных класса по числу особей были также одинаковыми (8,5%) и сумма их равна 17%.
Процент кроссинговера определяется как отношение числа гамет с зарегистрированными обменами между двумя определенными парами аллелей к общему числу гамет.
Значение частоты кроссинговера между двумя генами, выявляемое в опыте, не может быть более 50 %, т.к. эта частота составляет вероятность нормального, т.е. без кроссинговера, расхождения хромосом.