

Оксидазный путь использования кислорода в клетке - окислительное фосфорилирование
Оксидазный путь использования кислорода состоит из процессов окисления и фосфорилирования, которые между собой сопряжены. В нем участвует около 40 различных белков. Оксидазный путь потребляет 90% О2, является основным источником АТФ в аэробных клетках.
Окислительным фосфорилированием называют синтез АТФ из АДФ и Н3РО4 за счет энергии переноса электронов по ЦПЭ. При окислении выделяется 220 кДж/моль свободной энергии. На синтез 3 АТФ расходуется: 30,5*3=91,5 кДж/моль. В виде тепла выделяется: 220-91,5=128,5 кДж/моль. КПД = 40%.
НАДН2 + ½О2 → НАД+ + Н2О + 220 кДж/моль
АДФ + Н3РО4 + 30,5 кДж/моль = АТФ + Н2О
Компоненты цепи окислительного фосфорилирования
1). Цепь окисления (дыхательная цепь) состоит из 4 белковых комплексов, которые определенным образом встроены во внутреннюю мембрану митохондрий и небольших подвижных молекул убихинона и цитохрома С, которые циркулируют в липидном слое мембраны между белковыми комплексами.
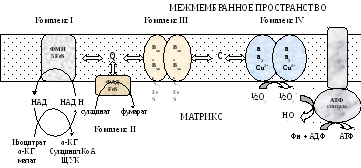
Комплекс I – НАДН2 дегидрогеназный комплекс – самый большой из дыхательных ферментных комплексов – имеет молекулярную массу свыше 800КДа, состоит из более 22 полипептидных цепей, в качестве коферментов содержит ФМН и 5 железо-серных (Fe2S2 и Fe4S4) белков.
Комплекс II – СДГ. В качестве коферментов содержит ФАД и железо-серный белок.
Комплекс III – Комплекс b-c1 (фермент QH2 ДГ), имеет молекулярную массу 500КДа, состоит из 8 полипептидных цепей, и вероятно существует в виде димера. Каждый мономер содержит 3 гема, связанных с цитохромами b562, b566, с1, и железо-серный белок.
Комплекс IV – Цитохромоксидазный комплекс имеет молекулярную массу 300КДа, состоит из 8 полипептидных цепей, существует в виде димера. Каждый мономер содержит 2 цитохрома (а и а3) и 2 атома меди.
Коэнзим Q (убихинон). Липид, радикал которого у млекопитающих образован 10 изопреноидными единицами (Q10). Убихинон переносит по 2Н+ и 2е-.
 убихинон
↔ семихинон
↔ гидрохинон
убихинон
↔ семихинон
↔ гидрохинон
Цитохром с. Периферический водорастворимый мембранный белок с массой 12,5КДа, содержит 1 полипептидную цепь из 100 АК, и молекулу гема.
Молекулярные соотношения между компонентами дыхательной цепи отличаются в разных тканях. Например, в миокарде, на 1 молекулу НАДН2 дегидрогеназного комплекса приходиться 3 молекулы комплекса b-c1, 7 молекул цитохромоксидазного комплекса, 9 молекул цитохрома С и 50 молекул убихинона.
2). Фосфорилирование осуществляется АТФ-синтетазой (Н+-АТФ-аза) — интегральным белком внутренней мембраны митохондрий. АТФ-синтаза состоит из 2 белковых комплексов, обозначаемых как F0 и F1. Гидрофобный комплекс F0 погружён в мембрану.
|
|
Он служит основанием, которое фиксирует АТФ-синтазу в мембране. Комплекс F0 состоит из нескольких субъединиц, образующих канал, по которому протоны переносятся в матрикс. Комплекс F1 выступает в митохондриальный матрикс. Он состоит из 9 субъединиц (3α, 3β, γ, δ, ε). Субъединицы α и β уложены попарно, образуя «головку»; между а- и β-субъединицами располагаются 3 активных центра, в которых происходит синтез АТФ; γ, δ, ε – субъединицы связывают комплекс F1, с F0. |
