

18. Компоненты дыхательной цепи и их характеристика. Фмн и фад-зависимые дегидрогеназы. Строение окисленной и восстановленной форм фмн.
Компоненты ДЦ
1 .НИКОТИНЗАВИСИМЫЕ ДГ, т.е. содержащие КОФЕРМЕНТЫ - НАД, НАДФ
2.ФЛАВИНЗАВИСИМЫЕ ДГ, т.е. содержащие КОФЕРМЕНТЫ - ФМН, ФАД.
З.УБИХИНОН (Ko-Q).
4.ЦИТОХРОМЫ: в, с, c1., а, а3.
Почти все эти компоненты, за исключением первого, встроены во внутреннюю мембрану МИТОХОНДРИЙ.
ФАД и ФМН-зависимые дегидрогеназы содержат в качестве кофермента фосфорный эфир витамина В2 (ФАД).
СТРОЕНИЕ КОМПОНЕНТОВ ДЫХАТЕЛЬНОЙ ЦЕПИ.
1 .В НАД и НАДФ рабочей частью является витамин РР - НИКОТИНАМИД.

НАД*2Н + 2е = НАДН+Н
2.В ФАД и ФМН рабочей частью является ФЛАВИИ (компонент витамина В2)

3.УБИХИНОН легко переходит в восстановленную форму KOQ +2Н +2е =KOQ*H2
4.ЦИТОХРОМЫ - это ГЕТЕРОПРОТЕИНЫ. Их белковой частью является ГЕМ. Белки хромопротеиды, способные присоединять электроны, благодаря наличию в своем составе в качестве простетических групп железопорфиринов. Они принимают электрон от вещества, являющегося немного боле сильным восстановителем, и передают его более сильному окислителю.
Цитохромы электронтранспортной цепи. Их функционирование. Образование воды как конечного продукта обмена.
ЦИТОХРОМЫ - это ГЕТЕРОПРОТЕИНЫ. Их белковой частью является ГЕМ, структура которого представляет собой 4 ПИРРОЛЬНЫХ кольца и атом железа, который легко меняет валентность. Также могут включать медь.

20. Пути синтеза атф. Субстратное фосфорилирование (примеры). Молекулярные механизмы окислительного фосфорилирования (теория Митчелла). Разобщение окисления и фосфорилирования.
Процесс образования АТФ в дыхательной цепи – окислительное фосфорилирование. За счет энергии транспорта электронов в ДЦ из АДФ и неорганического фосфата образуется АТФ. Субстратное фосфорилирование – процесс синтеза АТФ из АДФ и фосфата за счет энергии окисленного субстрата в цитоплазме клетки. Примером субстратного фосфорилирования могут служить реакции:

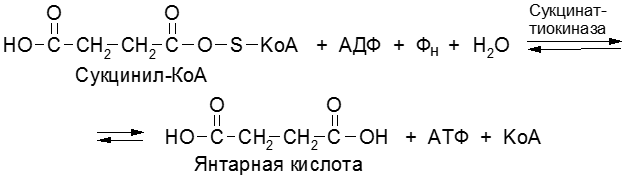
Основные положения теории Митчела:
1.Мембрана митохондрий не проницаема для протонов.
2.Образуется протонный потенциал в процессе транспорта электронов и протонов.
3.Обратный транспорт протонов в матрикс сопряжен с образованием атф.
Процесс транспорта электронов происходит во внутренней мембране. Протоны переносятся в межмембранное пространство, а электроны продвигаются по дыхательной цепи. Внутренняя мембрана со стороны матрикса заряжается отрицательно, а со стороны межмембранного пространства - положительно. Во время дыхания создается ЭЛЕКТРО-ХИМИЧЕСКИЙ градиент; концентрационный и разности потенциалов. Электрический и концентрационный градиент составляет ПРОТОНДВИЖУЩУЮ силу, которая дает силу для синтеза АТФ. На определенных участках внутренней мембраны есть протонные каналы. Протоны могут проходить обратно в матрицу, при этом образующаяся энергия идёт на синтез АТФ.
Разобщение дыхания и фосфорилирования
Некоторые химические вещества (протонофоры) могут переносить протоны или другие ионы (ионофоры) из межмембранного пространства через мембрану в матрикс, минуя протонные каналы АТФ-синтазы. В результате этого исчезает электрохимический потенциал и прекращается синтез АТФ. Это разобщение дыхания и фосфорилирования. В результате разобщения количество АТФ снижается, а АДФ увеличивается. Разобщители - липофильные вещества, легко проходящие через липидный слой мембраны. Это 2,4-динитрофенол, присоединяющий протон в межмембранном пространстве и перенося его в матрикс.