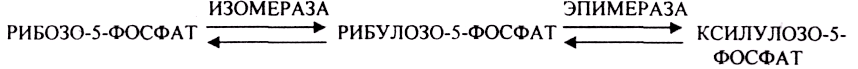42. Катаболизм глюкозы в анаэробных условиях. Химизм процесса, биологическая роль.
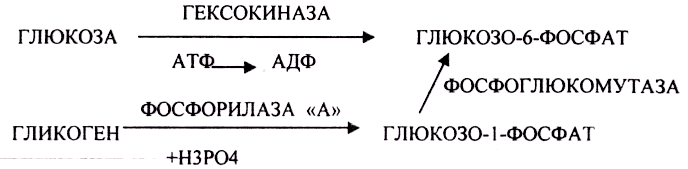
АНАЭРОБНЫЙ ГЛИКОЛИЗ протекает в цитоплазме клеток. Окисление глюкозы или глюкозного остатка гликогена всегда завершается образованием конечного продукта этого процесса- лактата. Окисление глюкозы и глюкозного остатка гликогена в тканях отличается начальных стадиях превращения, до образования глюкозо-6-фосфата. Дальнейшее окисление с этого этапа в тканях как в АНА-, так и в АЭРОБНЫХ условиях полностью совпадает до стадии образования ПИРУВАТА.
Процесс АНАЭРОБНОГО ГЛИКОЛИЗА сложный и многоступенчатый:
-первая стадия заканчивается образованием из ГЕКСОЗЫ двух ТРИОЗ -ДИОКСИАЦЕТОНФОСФАТА и ГЛИЦЕРАЛЬДЕГИД-3-ФОСФАТА.
-Вторая стадия - ГЛИКОЛИТИЧЕСКОЙ ОКСИДОРЕДУКЦИИ. Она сопряжена с образованием АТФ за счёт реакций СУБСТРАТНОГО ФОСФОРИЛИРОВАНИЯ, окислением ГЛИЦРАЛЬДЕГИД -3-ФОСФАТА, восстановлением ПИРУВАТА до ЛАКТАТА.
.В
процессе превращения глюкозы было
израсходовано 2 молекулы АТФ (ГЕКСОКИНАЗНАЯ
и ФОСФОФРУКТОКИНАЗНАЯ реакции). С этапа
образования ТРИОЗ идёт одновременное
их окисление. В результате этих реакций
образуется энергия в виде АТФ за счёт
реакций СУБСТРАТНОГО ФОСФОРИЛИРОВАНИЯ
(ГЛИЦЕРАТКИНАЗНАЯ и ПИРУВАТКИНАЗНАЯ).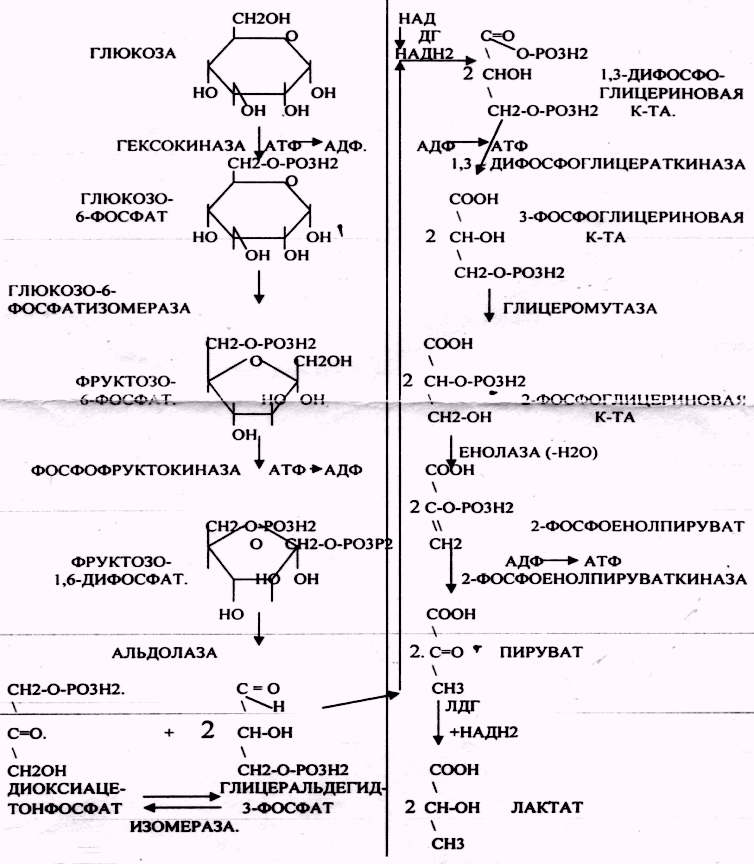
На этапе ГЛИКОЛИТИЧЕСКОЙ ОКСИДОРЕДУКЦИИ идёт окисление ГЛИЦЕРАЛЬДЕГИД-3-ФОСФАТА в присутствии НЗРО4 и НАД- зависимой ДГ. ПВК, принимая НАДН2, восстанавливается с образованием ЛАКТАТА.
43. Катаболизм глюкозы в тканях в аэробных условиях. Гексозодифосфатный путь превращения глюкозы и его биологическая роль. Эффект Пастера.
ГЕКСОЗОДИФОСФАТНЫЙ ПУТЬ - это аэробное превращение глюкозы в тканях. При поступлении кислорода в клетки происходит подавление анаэробного ГЛИКОЛИЗА. При этом понижается потребление глюкозы, блокируется образование ЛАКТАТА. Эффект торможения анаэробного гликолиза дыханием получил название эффекта ПАСТЕРА.
Процесс окисления начинается в цитоплазме до стадии образования ПИРУВАТА. Затем ПВК поступает в МИТОХОНДРИИ, где в матрице подвергается ОКИСЛИТЕЛЬНОМУ ДЕКАРБОКСИЛИРОВАНИЮ Образующийся АЦЕТИЛ-КОА поступает для дальнейшего окисления в основной метаболический ЦТК КРЕБСА. С участием ферментов ЦТК и сопряженных с ним ферментов дыхательной цепи происходит образование конечных продуктов (СО2 и Н2О) И выделяется 38 АТФ, а при окислении глюкозного остатка гликогена - 39 АТФ. Н2О образуется на этапе превращения:
ГЛИЦЕРАЛЬДЕГИД-3-ФОСФАТ
2-ФОСФОГЛИЦЕРИНОВАЯ К-ТА
ПИРУВАТА
Альфа- КЕТОГЛУТАРОВАЯ К-ТА
СУКЦИНАТ
ИЗОЦИТРАТ
МАЛАТ
СО2 образуется на этапе превращения;
ПИРУВАТ
ОКСАЛОСУКЦИНАТ
Альфа-КЕТОГЛУТАРОВАЯ К-ТА,
АТФ образуется:
А. За счет реакций субстратного ФОСФОРИЛИРОВАНИЯ на этапе превращения:
1,3-ДИФОСФОГЛИЦЕРИНОВАЯ К-ТА
2-ФОСФОЕНОЛПИРУВАТ
СУКЦИНИЛ-КОА
В. За счёт реакций ОКИСЛИТЕЛЬНОГО ФОСФОРИЛИРОВАНИЯ на этапе превращения:
ГЛИЦЕРАЛЬДЕГИД-3-ФОСФАТ
ПИРУВАТ
ИЗОЦИТРАТ
альфа-КГК
СУКЦИНАТ
МАЛАТ.
44. Гексозомонофосфатный путь превращения глюкозы в тканях и его биологическая роль.
Реакции окислительной стадии образования пентоз.
ГЕКСОЗОМОНОФОСФАТНЫЙ ПУТЬ протекает в цитоплазме клетки и представлен 2 ветвями: окислительной и неокислительной. Особенно активно этот путь протекает в тех органах и тканях, в которых активно синтезируются жиры. Биологическая роль этого пути окисления глюкозы анаболическая и связывается прежде всего с производством двух веществ:
1. НАДФ*Н2, который поступает в клетку для реакций синтеза и восстановления веществ.
2. РИБОЗО-5-ФОСФАТА и др. ПЕНТОЗ, которые используются в клетке для синтеза важнейших биологических молекул: ДНК, РНК, НТФ (АТФ, ГТФ, ЦТФ, ТТФ),Н5КОА, НАД, ФАД.
Неокислительная стадия ПЕНТОЗНОГО ПУТИ окисления глюкозы даёт субстраты, которые в анаэробных условиях поддерживают ГЛИКОЛИЗ (ФРУКТОЗО-6-ФОСФАТ, ГЛИЦЕРАЛЬДЕГИД-3-ФОСФАТ).
Окислительная стадия представляет 5 реакций и предусматривает образование ПЕНТОЗ. ГЕКСОЗОМОНОФОСФАТНЫЙ путь отличается от ГЕКСОЗОДИФОСФАТНОГО пути с этапа превращения глюкозо-6-фосфата.
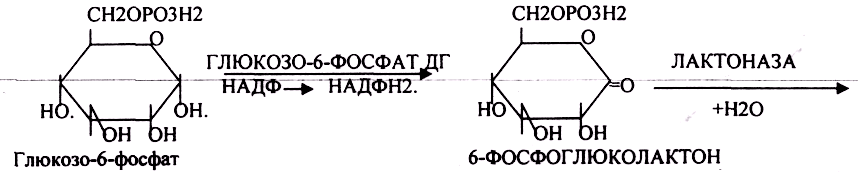


При определенных условиях на этом заканчивается окислительная стадия. Между ПЕНТОЗАМИ устанавливается подвижное равновесие.