

Pivovarennaya_inzheneria_ / Глава 3
.pdfДРОЖЖИ И ДРОЖЖЕВОЕ ХОЗЯЙСТВО ПИВОВАРЕННОГО ПРОИЗВОДСТВА |
105 |
|
|
Г л а в а 3
ДРОЖЖИ И ДРОЖЖЕВОЕ ХОЗЯЙСТВО ПИВОВАРЕННОГО ПРОИЗВОДСТВА
Роль дрожжей в процессе брожения была выявлена лишь в середине XIX в. Так, в средневековой версии баварской заповеди чистоты, принятой в 1516 г. по инициа тиве баварского герцога Вильгельма IV, о дрожжах вообще ничего не упоминалось. О них в то время попросту ничего не было известно. В тексте оригинала, этого по сути первого в мире закона о защите прав потребителя, буквально записано: «Мы хотим, чтобы во всех наших городах, на ярмарках и в деревнях не добавляли в пиво ничего, кроме ячменя, хмеля и воды».
А как же происходило брожение без внесения дрожжей? Приготовленное охлажден ное сусло оставляли в открытом сосуде, дрожжевые клетки (так называемые «дикие» дрож жи) попадали в него из воздуха и достаточно легко развивались в благоприятной среде — происходило так называемое самозабраживание. Помимо этого, для последующих цик лов брожения использовали осадки, образующиеся в бродильных емкостях. Кстати, при готовление пива самоброжением сохранялось в отдельных странах Европы, например Бельгии, вплоть до конца XIX в. При использовании «диких» дрожжей невозможно обес печить стабильность вкуса и полное освобождение пива от мути.
Антони ван Левенгук (1632–1675) впервые увидел в микроскоп дрожжи в 1680 г., однако лишь в 1830 г. эти образования признали живыми одноклеточными микроор ганизмами. В том же году Каньяром де Латуром, Кютцингом и Шваном было впер вые высказано предположение, что брожение обусловлено деятельностью микроор ганизмов, и в том числе дрожжей. Однако ученые того времени во главе с химиком Либихом вплоть до второй половины XIX в. противились этой гипотезе и, не пони мая сути происходящих явлений, считали, что в основе сбраживания сусла лежат химические процессы.
Биологическая сущность брожения и роль дрожжей, которую они играют в этом, была убедительно доказана Луи Пастером (1857), который, кстати, так и умер, пола гая, что активным началом брожения являются исключительно живые микроскопи ческие организмы — дрожжи. Впоследствии было показано, что и неживые биологи ческие объекты — ферменты — будучи выделенными из живой клетки, будут обладать теми же уникальными биокаталитическими способностями и так же возбуждать бро жение в отсутствие дрожжевых клеток.
В70 е гг. XIX в. на пивоварнях появились первые лаборатории, в которых начали осу ществлять систематические исследования дрожжей и выращивание чистой культуры, что после применения хмеля в пивоварении, пожалуй, явилось вторым революционным пре образованием в технологии пива.
3.1.ПИВНЫЕ ДРОЖЖИ
Внастоящем разделе приведены общие сведения о пивных дрожжах как об одном из основных объектов пивоварения, специфические особенности которого необходимо учи тывать при машинно аппаратурном обеспечении производства.
106 |
ПИВОВАРЕННАЯ ИНЖЕНЕРИЯ |
|
|
3.1.1. Характеристика пивных дрожжей
Пивные дрожжи представляют собой одноклеточные почкующиеся микроскопичес кие организмы округлой, овально яйцевидной формы. Основные параметры пивных дрож жей приведены в табл. 3.1.
Таблица 3.1
Геометрические и физические параметры дрожжевой клетки
Наименование |
Величина |
|
|
Ширина клетки, мкм |
5–8 |
Длина клетки, мкм |
8–11 |
Диаметр клетки, мкм |
6–10 |
Площадь поверхности клетки, м2 |
(172–200) 10–12 |
Объем клетки, м3 |
(150–400) 10–18 |
Масса клетки, кг |
(0,24–1,92) 10–13 |
Масса сухой клетки, кг |
Около 2,5 10–14 |
Плотность, кг/м3 |
1050–1189 |
Количество клеток в 1 г СВ |
Около 4 10–10 |
Техническое значение дрожжей обусловлено их специфической способностью пре вращать сахар в этиловый спирт и диоксид углерода. По этой причине они издавна получили общее название сахаромицетов или сахарных грибов — Saccharomyces.
3.1.2. Виды пивных дрожжей
Многообразие штаммов пивных дрожжей составляет две группы — дрожжи низового и верхового брожения.
Дрожжи низового брожения в большинстве представляют собой отдельные клетки или их пары, тогда как дрожжи верхового брожения образуют клеточные сообщества. Это обусловлено тем, что материнские и дочерние клетки дрожжей низового броже ния после деления сразу отделяются друг от друга, в то время как у дрожжей верхового брожения материнская и дочерняя клетки долго связаны между собой, вследствие чего образуются разветвленные сообщества клеток.
К одному из физиологических различий дрожжей низового и верхового броже ния относят их способность к сбраживанию рафинозы, представляющей собой триса харид — углевод, состоящий из трех остатков моносахаридов, связанных гликозидны ми связями. Набор ферментов низовых дрожжей (хотя он беднее, чем у верховых) обеспечивает полную переработку рафинозы, а верховые дрожжи сбраживают ее только на треть.
Помимо этого дрожжи низового и верхового брожения различаются особенностя ми обмена веществ и способностью к спорообразованию. Если у дрожжей низового
ДРОЖЖИ И ДРОЖЖЕВОЕ ХОЗЯЙСТВО ПИВОВАРЕННОГО ПРОИЗВОДСТВА |
107 |
|
|
брожения обмен веществ основан преимущественно на брожении, то у дрожжей вер хового брожения — в значительной степени на дыхании. Этим обусловлен бóльший выход биомассы у дрожжей верхового брожения.
Дрожжи низового брожения реже образуют споры, а спорообразование у них продол жительнее.
3.1.3. Морфология дрожжей
Строение клеток дрожжей низового и верхового брожения одинаково (рис. 3.1) — они состоят из протопласта и оболочки.
7 |
8 |
|
9 |
10 |
|
11 |
6 |
|
|
|
|
|
|
5 |
|
|
|
|
|
12 |
|
|
|
|
|
|
|
4 |
|
|
|
|
|
|
3 |
|
|
|
|
|
13 |
|
|
|
|
|
|
|
2 |
|
|
|
|
|
14 |
|
|
|
|
|
|
|
1 |
|
|
|
|
|
15 |
|
|
|
|
|
|
|
0 |
1 |
2 |
3 |
4 |
5 |
мкм |
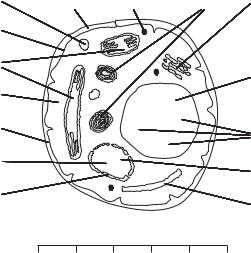
Рис. 3.1. Строение дрожжевой клетки:
1 — мембрана ядра; 2 — ядрышко; 3 — клеточная оболочка; 4 — цитоплазма; 5 — митохондрии;
6 — цитоплазматическая мембрана; 7 — рибосомы;
8 — рубец; 9 — гранулы резервных веществ;
10 — липидная гранула; 11 — комплекс Гольджи;
12 — вакуоль; 13 — полиметафосфатная гранула;
14 — клеточное ядро; 15 — эндоплазматическая сеть
В протопласте дрожжей различают цитоплазматическую мембрану 6, расположенную непосредственно под оболочкой 3, цитоплазму 4 — вязкую жидкость со структурными элементами (рибосомами 7, митохондриями 5) и ядро 14. Еще в цитоплазме содержатся включения запасных питательных веществ в виде капель жира, а также гранулы 9 гликоге на и волютина. По мере роста дрожжевых клеток в них появляются вакуоли 12 — полости, заполненные клеточным соком. Рубец 8 — место, где отпочковалась дочерняя клетка.
108 |
ПИВОВАРЕННАЯ ИНЖЕНЕРИЯ |
|
|
Характеристика основных структурных элементов клетки приведена в табл. 3.2.
|
|
|
Таблица 3.2 |
|
Структурные элементы клетки и их характеристики |
||
|
|
|
|
Структурные |
|
Функциональное назначение |
Основные компоненты |
элементы |
|
|
структурного элемента |
Клеточная облочка |
|
Механический барьер, массоперенос |
Полисахариды, белки |
|
|
|
липиды и др. |
Цитоплазматическая |
|
Селективный массоперенос |
Мембраны, ферменты |
мембрана |
|
|
|
Ядро |
|
Хранение генетической информации, |
ДНК, белки и ферменты, |
|
|
репликация ДНК, образование инA |
связанные с образованиA |
|
|
формационных РНК (транскрипция) |
ем ДНК и РНК |
Митохондрии |
|
Энергетический центр клетки, обраA |
Мембраны, ферменты |
|
|
зование АТФ, дыхание, окисление |
цикла Кребса и дыхаA |
|
|
питательных веществ и др. |
тельной цепи |
Рибосомы |
|
Синтез белка |
РНК, белки |
Лизосомы |
|
Разрушение биополимеров — трансA |
Мембраны, ферменты |
|
|
формация высокомолекулярных соA |
|
|
|
единений в низкомолекулярные |
|
Эндоплазматическая |
|
Синтез липидов и других веществ |
То же |
сеть |
|
|
|
Комплекс Гольджи |
|
Экскреция — удаление токсичных веществ |
— “ — |
Вакуоли |
|
Накопление резервных и ненужных |
— ” — |
|
|
клетке веществ |
|
Гранулы |
|
Резервные вещества |
Гликоген, волютин и др. |
Клеточная оболочка представляет собой основной структурный элемент клет ки, состоящий на 70% из полисахаридов; остальное составляют белки, липиды и пр. Функциональная роль клеточной оболочки — механический барьер, обеспе чивающий форму клетки, поддержание осмотического давления в ней и участие
втранспорте веществ.
Уодних видов дрожжей оболочки могут в той или иной степени ослизняться, вследствие чего клетки к окончанию брожения, склеиваясь между собой и образуя хлопья, оседают на дно бродильного аппарата. Такие дрожжи называют хлопьевидными. К ним, в частности, относят пивные дрожжи низового брожения (Saccharomyces carlsbergensis).
Удругих видов дрожжей оболочки клеток не ослизняются и клетки не слипаются друг с другом. Такие дрожжи называют пылевидными. К ним относят дрожжи верхового брожения (Saccharomyces cerevisae), которые при брожении всплывают на поверхность и накапливаются в густом слое пены (последнюю при открытом способе брожения периодически удаляют). В закрытых бродильных аппаратах по окончании брожения пылевидные дрожжи также оседают, но образуют рыхлый осадок.
Цитоплазматическая мембрана расположена непосредственно под клеточной обо лочкой, имеет толщину около 8 нм и состоит из белков и липоидов. Функциональная роль мембраны — обеспечение селективного массопереноса, благодаря которому про исходит регулирование обмена веществ клетки.
ДРОЖЖИ И ДРОЖЖЕВОЕ ХОЗЯЙСТВО ПИВОВАРЕННОГО ПРОИЗВОДСТВА |
109 |
|
|
Рибосомы — небольшие гранулы, содержащие 58% белка и 42% рибонуклеиновой кислоты (РНК). Функциональная роль рибосом — осуществление процессов, направ ленных на синтез белка.
Митохондрии — небольшие гранулы или ветвистые нити, содержащие белки, ли пиды, фосфолипиды, жирные кислоты, ферменты и дезоксирибонуклеиновую кисло ту (ДНК). Функциональная роль митохондрий — осуществление процессов, направ ленных на обеспечение клетки энергией.
Ядро — обособленный от цитоплазмы органоид, окруженный собственной оболоч кой. Ядро состоит в основном из хроматина, регулирующего обмен веществ, синтез белка, передачу наследственных признаков.
Вакуоли — пузырьки, заполненные клеточным соком, содержащим соли металлов, углеводы, некоторые простые белки и жиры. В вакуолях накапливаются резервные вещества, среди которых гликоген и волютин. Гликоген — углевод, накапливающийся в клетках при брожении в условиях обильного углеводного питания. При ограничении питания и интенсивной аэрации, быстром росте и размножении клеток гликоген исче зает. Волютин присутствует в дрожжевых клетках, культивируемых в средах, богатых фосфатами.
Величина вакуолей зависит от возраста клеток и от условий культивирования. В молодых клетках вакуоли незаметны, а в старых или угнетенных клетках они могут занимать до 80% объема.
3.1.4. Химический состав дрожжей
Прессованные дрожжи содержат 65–75% воды, из которой около 60% представляет собой внутриклеточную влагу. В сухом веществе дрожжей содержится: 40–60% соеди нений азота, 25–35% углеводов, 4–7% жиров (липидов), 6–9% минеральных веществ, ферменты, витамины и пр.
Среди минеральных веществ доминируют (ориентировочно на 100 г сухого вещества): калий — 2400 мг, фосфаты — 2000 мг, натрий — 200 мг, кальций — 20 мг, магний — 2 мг и, кроме того, около 7 мг прочих микроэлементов (цинк, следы железа, марганца и меди).
Ферменты дрожжей сосредоточены в основном в клеточных мембранах, цитоплаз ме, вакуолях и ядре клетки. Ферменты способствуют усвоению питательных веществ, обмену веществ, росту и размножению клеток.
Содержание витаминов в дрожжах (на 100 г сухого вещества): тиамин (В1) — 8–15 мг; рибофлавин — 2–8 мг; никотиновая кислота — 30–100 мг; фолиевая кислота — 2–10 мг; пантотеновая кислота — 2–20 мг; пиридоксин — 3–10 мг; биотин — 0,1–1 мг.
3.2. ТЕХНОЛОГИЧЕСКИЕ АСПЕКТЫ КУЛЬТИВИРОВАНИЯ МИКРООРГАНИЗМОВ
Дрожжи, как живые организмы, являются важнейшим «партнером» пивовара, причем жизненные «интересы» дрожжевой клетки диаметрально противоположны производствен ным интересам пивовара. Так, пивовар в первую очередь заинтересован в образовании
110 |
ПИВОВАРЕННАЯ ИНЖЕНЕРИЯ |
|
|
спирта и диоксида углерода при сбраживании сусла, в то время как для дрожжей эти продукты их жизнедеятельности токсичны, и поэтому они стремятся избавиться от них, выделяя наружу.
Для дрожжевой клетки имеет значение лишь потребление питательных веществ с целью получения энергии для продолжения своего существования и образования клеточной массы.
Поскольку качество пива в значительной степени зависит от дрожжей и их жизне деятельности, то роль пивовара при брожении сводится к тому, чтобы обеспечить оп тимальные условия для жизнедеятельности дрожжей, целенаправленно и эффективно управляя факторами, которые регулируют обмен веществ дрожжей.
3.2.1. Обмен веществ дрожжевой клетки
Материальную основу роста и размножения клеток составляет ассимиляция ве ществ из окружающей среды, так называемый конструктивный («строительный») об мен — анаболизм.
Конструктивный обмен требует определенного расхода энергии и невозможен без энер гетического обмена — катаболизма.
Клетка осуществляет комплексный обмен веществ — углеводный, белковый, липид ный и обмен минеральных веществ.
Энергию, необходимую для поддержания жизнедеятельности и роста, микроорганиз мы получают в результате углеводного обмена. Различают аэробные и анаэробные мик роорганизмы. Первые получают энергию при окислении питательного субстрата в про цессе дыхания, а вторые — в результате брожения.
Дрожжи — единственный живой организм, способный при недостатке воздуха перехо дить с аэробного, энергетически более выгодного обмена веществ, на анаэробный.
Поскольку в дрожжевой клетке процессы брожения и дыхания взаимосвязаны, то фер ментативные процессы в ней можно направить либо в сторону брожения, способствую щего образованию спирта, либо в сторону дыхания, способствующего накоплению био массы. Направленность процесса обусловлена количеством кислорода, растворенного в культуральной среде.
При наличии кислорода в среде дрожжи в процессе дыхания полностью расщепляют
глюкозу с образованием воды и диоксида углерода: |
|
С6Н12О6 + 6О2 → 6Н2О + 6СО2 + 28 200 кДж. |
(3.1) |
При отсутствии воздуха дрожжи переходят на спиртовое брожение, в результате кото
рого образуется этанол и диоксид углерода |
|
С6Н12О6 → 2С2Н5ОН+2СО2+1130 кДж. |
(3.2) |
Биотрансформация глюкозы в дрожжевой клетке осуществляется путем большого числа последовательных реакций, каждая из которых катализируется специфическим ферментом. В дрожжевой клетке эти ферменты связаны с определенными клеточными
ДРОЖЖИ И ДРОЖЖЕВОЕ ХОЗЯЙСТВО ПИВОВАРЕННОГО ПРОИЗВОДСТВА |
111 |
|
|
структурами. Например, ферменты для обеспечения спиртового брожения находятся в цитоплазме, а ферменты для обеспечения дыхания локализованы в митохондриях.
Жизнедеятельность дрожжей в аэробных условиях характеризуется:
•получением большего количества энергии;
•хорошим физиологическим состоянием клеток;
•полной биодеградацией углеводов до воды и диоксида углерода. Жизнедеятельность дрожжей в анаэробных условиях сопровождается:
•получением существенно меньшего количества энергии;
•ослаблением физиологического состояния клеток;
•неполной биодеградацией углеводов до спирта и диоксида углерода. Органические вещества, необходимые клеткам для получения энергии, избиратель
но транспортируются внутрь клетки через клеточную мембрану. При этом селектив ность этого массопереноса обеспечивается специфическими транспортными механиз мами, зависящими от количества и типов ферментов дрожжевой клетки.
Белковый обмен веществ, так же, как липидный и обмен минеральных веществ, направлен прежде всего на образование новых клеточных субстанций. При этом суще ственную роль играют не только процессы синтеза, но и процессы распада.
3.2.2. Основы кинетики развития микроорганизмов
Настоящий раздел посвящен преимущественно биотехнологии чистой культуры дрожжей. Но поскольку на современных пивоваренных предприятиях стали повсемест но производить для собственных технологических нужд молочную кислоту, продуциру емую молочнокислыми бактериями, то технологические основы культивирования микро организмов изложены в общем виде (без разделения биологических объектов на дрожжи
ибактерии). Затем будут описаны практические способы и оборудование только для получения чистых дрожжевых культур, а установки для получения молочной кислоты описаны в главе 9, посвященной вспомогательному оборудованию.
Аппараты, в которых осуществляют культивирование микроорганизмов, называют фер ментерами, ферментаторами, дрожжегенераторами, пропагаторами и пр. Будем называть их единым термином, применяемым в последнее время наиболее широко, — биореакторы.
Вбиореакторах обеспечивают оптимальные условия для жизнедеятельности микро организмов, при этом в клетках осуществляется биосинтез, в результате которого в ре акционном объеме происходит накопление биомассы и продуктов метаболизма. В од них случаях целевым продуктом является биомасса, как, например, при получении чистой культуры дрожжей, а в других — прочие продукты биосинтеза, в том числе метаболиты, например молочная кислота и пр.
Воснову технологических расчетов оборудования для культивирования микроор ганизмов положена кинетика — учение о скорости протекания процессов.
Эффективная реализация культивирования микроорганизмов, помимо физиологиче ских свойств культивируемого штамма, зависит от конструктивного устройства био реактора, структуры потоков и их гидродинамических режимов, скоростей массо
итеплопереноса и пр. Совокупное влияние этих факторов определяет так называемую макрокинетику, описывающую развитие микроорганизмов не в идеальных, а в реаль ных условиях в большом объеме — в аппарате, в котором развиваются миллиарды
112 |
ПИВОВАРЕННАЯ ИНЖЕНЕРИЯ |
|
|
микроорганизмов при массовом движении макрочастиц — образований микроорга низмов, частиц субстрата, капель, струй, пузырей и т. п.
При изучении макрокинетики используют данные по микрокинетике, описываю щей элементарные процессы, протекающие на молекулярном уровне независимо друг от друга, с учетом предположения об упрощенном механизме их осуществления. К та ким процессам относят, например, молекулярную диффузию, теплопроводность, об мен веществ биологического объекта (усвоение им продуктов внешней среды и выде ление продуктов метаболизма) и т. д.
3.2.2.1.Скорость роста микроорганизмов
Впроцессе жизнедеятельности клетки можно выделить три этапа: рост, развитие
иделение. Рост клетки сопровождается увеличением биомассы организма за счет ус воения им питательных веществ среды. Развитие клетки характеризуется изменением морфологических и физиологических свойств в процессе жизненного цикла.
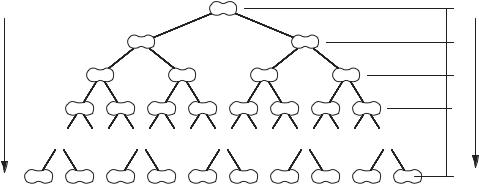
Если предположить, что условия внешней среды неизменны и все клетки делятся одновременно (синхронно), то процесс наращивания биомассы дрожжей можно пред ставить в виде следующей феноменологической модели (рис. 3.2).
τ0 = 0
... |
... |
... |
... |
... |
τ
Рис. 3.2. Схема размножения дрожжей
m0
ng g g g
m
Среднюю продолжительность жизни одного поколения (генерации) — промежу ток времени между двумя последовательными делениями клетки ∆t — называют вре менем генерации g, которое зависит как от внешних условий среды обитания, то есть от параметров культивирования, так и от штамма и вида микроорганизма, и может состав лять от нескольких минут до нескольких часов, например, для бактериальной клетки g = 20–30 мин, а для животной клетки — около 24 ч. Для клетки пивных дрожжей время генерации в производственных условиях составляет, как правило, 6–9 часов и зависит от температуры (табл. 3.3).
ДРОЖЖИ И ДРОЖЖЕВОЕ ХОЗЯЙСТВО ПИВОВАРЕННОГО ПРОИЗВОДСТВА |
113 |
|
|
Таблица 3.3
Зависимость времени генерации дрожжей Saccharomyces cerevisiae от температуры
Температура, °С |
12 |
13 |
15 |
20 |
Время генерации g, ч |
12 |
11 |
8 |
5 |
Среднее число делений клетки за единицу времени: ψ = 1/g.
Вблагоприятных условиях развития популяции в неограниченных пространстве
иресурсах питания прирост биомассы ∆m (кг) пропорционален промежутку времени культивирования ∆t и исходному количеству биомассы m, которое находилось в био реакторе к началу упомянутого промежутка времени:
∆m = µ m ∆t, |
(3.3) |
где µ — коэффициент пропорциональности, ч –1.
Малым приращениям ∆τсоответствуют малые приращения ∆m, поэтому функциональ ную зависимость m = f(τ) можно считать непрерывной, что позволяет представить урав нение роста популяции микроорганизмов в дифференциальной форме
dm = µ m dt. |
(3.4) |
Преобразуя уравнение (3.4) делением левой и правой части на dτ, получим выра жение валовой, или общей скорости роста биомассы (кг/ч):
dm |
=µm, |
(3.5) |
|
dτ |
|||
|
|
где dm — прирост биомассы за бесконечно малый промежуток времени dτ.
3.2.2.2. Удельная скорость роста микроорганизмов
Дальнейшее преобразование уравнения (3.5) делением обеих его частей на m по зволяет получить выражение коэффициента пропорциональности µ, который называ ют коэффициентом скорости роста (ч –1):
µ = |
dm |
. |
(3.6) |
|
|||
|
mdτ |
|
|
Отсюда видно, что этот коэффициент пропорциональности по физической сущности представляет собой относительную, или удельную скорость роста микроорганиз мов, соответствующую приросту каждой единицы биомассы популяции за бесконечно малый промежуток времени.
114 |
ПИВОВАРЕННАЯ ИНЖЕНЕРИЯ |
|
|
Удельная скорость роста является комплексным показателем развития микробной популяции, учитывающим влияние разнообразных факторов. В связи с этим в биоре акторах необходимо обеспечивать такие условия культивирования, при которых удель ная скорость роста будет наиболее приближена к своему максимально возможному значению µmax, которое для каждой культуры строго индивидуально и постоянно, то есть является кинетической константой, устанавливаемой экспериментально. Это максимальное значение µmax является пределом, к которому стремится удельная скорость роста и который в реальных производственных условиях практически недостижим. Реальные значения удельной скорости роста в промышленных условиях обычно со ставляют 0,8–0,9 от максимально возможной величины.
3.2.2.3. Экспоненциальная закономерность роста микроорганизмов
Поделив левую и правую часть уравнения (3.4) на m, можно его представить в виде
dm |
= µdτ. |
(3.7) |
m |
|
|
Рассмотрим два состояния системы (рис. 3.2): начальное, характеризующееся на личием в биореакторе биомассы в количестве m0, и конечное — через промежуток времени τ, характеризующееся увеличением количества биомассы до m.
Проинтегрировав левую часть уравнения 3.7 от m0 до m, а правую — от 0 до τ, полу чим уравнение роста микроорганизмов, представляющее собой экспоненциальную за висимость
m = m |
0 |
еµτ, |
(3.8) |
где е — основание натурального логарифма, е = 2,718.
Аналогично можно вывести уравнения роста концентрации биомассы х (кг/м3) (3.9) и количества клеток N (шт.) (3.10):
m, кг
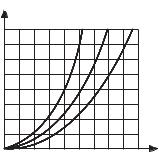
µ1 > µ2 > µ3 |
τ, ч |
Рис. 3.3 |
х = х |
0 |
еµτ, |
(3.9) |
N = N еµτ. |
(3.10) |
||
|
|
0 |
|
С математической точки зрения функция, представ ленная уравнением (3.10), непрерывна, поскольку вре мя непрерывно, а с физической точки зрения она диск ретна, поскольку количество микроорганизмов может быть только целым числом.
Таким образом, из вышеприведенных уравнений следует, что фаза бурного развития микроорганизмов в благоприятных условиях описывается экспоненци альной зависимостью. Графическая интерпретация этой зависимости приведена на рис. 3.3.
Однако в процессе жизнедеятельности микроорга низмов питательная среда истощается, а образовавшиеся