

Анаболизм и катаболизм
Анаболизм и катаболизм – это основные метаболические процессы.
Катаболизм – это ферментативное расщепление сложных органических соединений, осуществляющееся внутри клетки за счет реакций окисления. Катаболизм сопровождается выделением энергии и запасанием ее в макроэргических фосфатных связях АТФ.
Анаболизм – это синтез сложных органических соединений – белков, нуклеиновых кислот, полисахаридов – из простых предшественников, поступающих в клетку из окружающей среды или образующихся в процессе катаболизма. Процессы синтеза связаны с потреблением свободной энергии, которая поставляется АТФ (рис. 31).
![]()
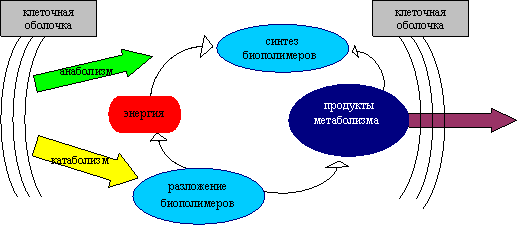
Рис. 31 Схема путей метаболизма в бактериальной клетке
В зависимости от биохимии процесса диссимиляции (катаболизма) различают дыхание и брожение.
Дыхание– это сложный процесс биологического окисления различных соединений), сопряженный с образованием большого количества энергии, аккумулируемой в виде макроэргических связей в структуре АТФ (аденозинтрифосфат), УТФ (уридинтрифосфат) и т.д., и образованием углекислого газа и воды. Различают аэробное и анаэробное дыхание.
Брожение– неполный распад органических соединений с образованием незначительного количества энергии и продуктов, богатых энергией.
Анаболизм включает процессы синтеза, при которых используется энергия, вырабатываемая в процессе катаболизма. В живой клетке одновременно и непрерывно протекают процессы катаболизма и анаболизма. Многие реакции и промежуточные продукты являются для них общими.
Живые организмы классифицируют в соответствии с тем, какой источник энергии или углерода они используют. Углерод – основной элемент живой материи. В конструктивном метаболизме ему принадлежит ведущая роль.
В зависимости от источника клеточного углерода все организмы, включая прокариотные, делят на автотрофы и гетеротрофы.
Автотрофыиспользуют CO2 в качестве единственного источника углерода, восстанавливая его водородом, который отщепляется от воды или другого вещества. Органические вещества они синтезируют из простых неорганических соединений в процессе фото- или хемосинтеза.
Гетеротрофы получают углерод из органических соединений.
Живые организмы могут использовать световую или химическую энергию. Организмы, живущие за счет энергии света, называют фототрофными.Органические вещества они синтезируют, поглощая электромагнитное излучение Солнца (свет). К ним относятся растения, сине-зеленые водоросли, зеленые и пурпурные серобактерии.
Организмы, получающие энергию из субстратов, источников питания (энергия окисления неорганических веществ), называют хемотрофами.Кхемогетеротрофамотносятся большинство бактерий, а так же грибы и животные.
Существует немногочисленная группа хемоавтотрофов. К таким хемосинтезирующим микроорганизмам относятся нитрифицирующие бактерии, которые, окисляя аммиак до азотистой кислоты, высвобождают необходимую для синтеза энергию. К хемосинтетикам относятся также водородные бактерии, получающие энергию в процессе окисления молекулярного водорода.
Углеводы как источник энергии
У большинства организмов расщепление органических веществ происходит в присутствии кислорода – аэробный обмен. В результате такого обмена остаются бедные энергией конечные продукты (СО2и Н2О), но высвобождается много энергии. Процесс аэробного обмена называется дыханием, анаэробного – брожением.
Углеводы – основной энергетический материал, который клетки используют в первую очередь для получения химической энергии. Кроме того, при дыхании могут использоваться также белки и жиры, а при брожении – спирты и органические кислоты.
Расщепление углеводов организмы осуществляют разными путями, в которых важнейшим промежуточным продуктом является пировиноградная кислота (пируват). Пируват занимает центральное место в метаболизме при дыхании и брожении. Выделяют три основных механизма образования ПВК.
1.Фруктозодифосфатный (гликолиз) или путь Эмбдена-Мейергофа-Парнаса– универсальный путь.
Процесс начинается с фосфорилирования (рис. 32). При участии фермента гексокиназы и АТФ глюкоза фосфорилируется по шестому углеродному атому с образованием глюкозо-6-фосфата. Это активная форма глюкозы. Она служит исходным продуктом при расщеплении углеводов любым из трех путей.
При гликолизе глюкозо-6-фосфат изомеризуется во фруктозо-6-фосфат, а затем под действием 6-фосфофруктокиназы фосфорилируется по первому углеродному атому. Образовавшийся фруктозо-1,6-дифосфат под действием фермента альдолазы легко распадается на две триозы: фосфоглицериновый альдегид и дигидроксиацетонфосфат. Дальнейшее превращение С3-углеводов осуществляется за счет переноса водорода и фосфорных остатков через ряд органических кислот с участием специфических дегидрогеназ. Все реакции этого пути, за исключением трех, протекающих с участием гексокиназы, 6-фосфофруктокиназы и пируваткиназы, полностью обратимы. На стадии образования пировиноградной кислоты заканчивается анаэробная фаза превращения углеводов.
Баланс:
![]()
Максимальное количество энергии, получаемое клеткой при окислении одной молекулы углеводов гликолитическим путем, равно 2·105Дж.
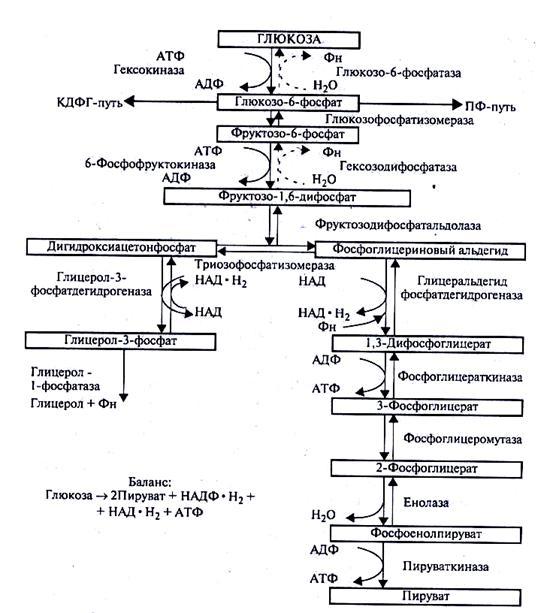
Рис.32. Фруктозодифосфатный путь расщепления глюкозы
2.Пентозофосфатный (Варбурга-Дикенса-Хорекера)путь характерен также для большинства организмов (в большей степени для растений, а для микроорганизмов играет вспомогательную роль). В отличие от гликолиза ПФ путь не образует пируват.
Глюкозо-6-фосфат превращается в 6-фосфоглюколактон, который декарбоксилируется (рис. 33). При этом образуется рибулозо-5-фосфат, на котором завершается процесс окисления. Последующие реакции рассматриваются как процессы превращения пентозофосфатов в гексозофосфаты и обратно, т.е. образуется цикл. Считают, что пентозофосфатный путь на одном из этапов переходит в гликолиз.
При прохождении через ПФ путь каждых шести молекул глюкозы происходит полное окисление одной молекулы глюкозо-6-фосфата до CO2и восстановление 6 молекул НАДФ+до НАДФ·Н2. Как механизм получения энергии этот путь в два раза менее эффективен, чем гликолитический: на каждую молекулу глюкозы образуется 1 молекула АТФ.

Рис. 33. Пентозофосфатный путь расщепления глюкозо-6-фосфата
Основное назначение этого пути – поставлять пентозы, необходимые для синтеза нуклеиновых кислот, и обеспечивать образование большей части НАДФ·Н2, необходимого для синтеза жирных кислот, стероидов.
3.Путь Энтнера-Дудорова (кетодезоксифосфоглюконатный или КДФГ-путь)встречается только у бактерий. Глюкоза фосфорилируется молекулой АТФ при участии фермента гексокиназы (рис. 34).
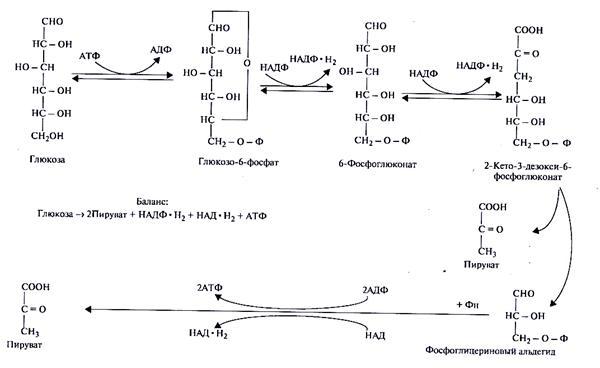
Рис.34. Путь Энтнера-Дудорова расщепления глюкозы
Продукт фосфорилирования – глюкозо-6-фосфат – дегидрируется до 6-фосфоглюконата. Под действием фермента фосфоглюконатдегидрогеназы от него отщепляется вода и образуется 2-кето-3-дезокси-6-фосфоглюконат (КДФГ). Последний расщепляется специфичной альдолазой на пируват и глицеральдегид-3-фосфат. Глицеральдегид далее подвергается действию ферментов гликолитического пути и трансформируется во вторую молекулу пирувата. Кроме того, этот путь поставляет клетке 1 молекулу АТФ и 2 молекулы НАД·Н2.
Таким образом, основным промежуточным продуктом окислительного расщепления углеводов является пировиноградная кислота, которая при участии ферментов превращается в различные вещества. Образовавшаяся одним из путей ПВК в клетке подвергается дальнейшему окислению. Освобождающиеся углерод и водород удаляются из клетки. Углерод выделяется в форме CO2, водород передается на различные акцепторы. Причем может передаваться либо ион водорода, либо электрон, поэтому перенос водорода равноценен переносу электрона. В зависимости от конечного акцептора водорода (электрона) различают аэробное дыхание, анаэробное дыхание и брожение.
Дыхание
Дыхание – окислительно-восстановительный процесс, идущий с образованием АТФ; роль доноров водорода (электронов) в нем играют органические или неорганические соединения, акцепторами водорода (электронов) в большинстве случаев служат неорганические соединения.
Если конечный акцептор электронов – молекулярный кислород, дыхательный процесс называют аэробным дыханием. У некоторых микроорганизмов конечным акцептором электронов служат такие соединения, как нитраты, сульфаты и карбонаты. Этот процесс называется анаэробным дыханием.
Аэробное дыхание – процесс полного окисления субстратов до CO2 и Н2О с образованием большого количества энергии в форме АТФ.
Полное окисление пировиноградной кислоты происходит в аэробных условиях в цикле трикарбоновых кислот (ЦТК или цикл Кребса) и дыхательной цепи.
Аэробное дыхание состоит из двух фаз:
1). Образующийся в процессе гликолиза пируват окисляется до ацетил-КоА, а затем до CO2, а освобождающиеся атомы водорода перемещаются к акцепторам. Так осуществляется ЦТК.
2). Атомы водорода, отщепленные дегидрогеназами, акцептируются коферментами анаэробных и аэробных дегидрогеназ. Затем они переносятся по дыхательной цепи, на отдельных участках которой образуется значительное количество свободной энергии в виде высокоэнергетических фосфатов.
Цикл трикарбоновых кислот (цикл Кребса, ЦТК)
Пируват, образующийся в процессе гликолиза, при участии мультиферментного комплекса пируватдегидрогеназы декарбоксилируется до ацетальдегида. Ацетальдегид, соединяясь с коферментом одного из окислительных ферментов – коферментом А (КоА-SH), образует «активированную уксусную кислоту» - ацетил-КоА – высокоэнергетическое соединение.

Ацетил-КоА под действием цитрат-синтетазы вступает в реакцию со щавелевоуксусной кислотой (оксалоацетат), образуя лимонную кислоту (цитрат С6), которая является основным звеном ЦТК (рис. 35). Цитрат после изомеризации превращается в изоцитрат. Затем следует окислительное (отщепление Н) декарбоксилирование (отщепление СО2) изоцитрата, продуктом которого является 2-оксоглутарат (С5). Под влиянием ферментного комплекса ɑ-кетоглутаратдегидрогеназы с активной группой НАД он превращается в сукцинат, теряя СО2 и два атома водорода. Сукцинат затем окисляется в фумарат (С4), а последний гидратируется (присоединение Н2О) в малат. В завершающей цикл Кребса реакции происходит окисление малата, что приводит к регенерации оксалоацетата (С4). Оксалоацетат взаимодействует с ацетил-КоА, и цикл повторяется снова. Каждая из 10 реакций ЦТК, за исключением одной, легко обратима. В цикл вступают два атома углерода в виде ацетил-КоА и такое же количество атомов углерода покидают этот цикл в виде СО2.
![]()



Рис. 35. Цикл Кребса (по В.Л. Кретовичу):
1, 6 – система окислительного декарбоксилирования; 2 – цитратсинтетаза, кофермент А; 3, 4 – аконитатгидратаза; 5 – изоцитратдегидрогеназа; 7 – сукцинатдегидрогеназа; 8 – фумаратгидратаза; 9 – малатдегидрогеназа; 10 – спонтанное превращение; 11 - пируваткарбоксилаза
В результате четырех окислительно-восстановительных реакций цикла Кребса осуществляется перенос трех пар электронов на НАД и одной пары электронов на ФАД. Восстановленные таким путем переносчики электронов НАД и ФАД подвергаются затем окислению уже в цепи переноса электронов. В цикле образуется одна молекула АТФ, 2 молекулы СО2 и 8 атомов водорода.
Биологическое значение цикла Кребса заключается в том, что он является мощным поставщиком энергии и «строительных блоков» для биосинтетических процессов. Цикл Кребса действует только в аэробных условиях, в анаэробных он разомкнут на уровне α-кетоглутаратдегидрогеназы.
Дыхательная цепь
Последней стадией катаболизма является окислительное фосфорилирование. В ходе этого процесса высвобождается большая часть метаболической энергии.
Восстановленные в цикле Кребса переносчики электронов НАД и ФАД подвергаются окислению в дыхательной цепи или цепи транспорта электронов. Молекулы-переносчики – это дегидрогеназы, хиноны и цитохромы.
Обе ферментные системы у прокариот находятся в плазматической мембране, а у эукариот – во внутренней мембране митохондрий. Электроны от атомов водорода (НАД, ФАД) по сложной цепи переносчиков переходят к молекулярному кислороду, восстанавливая его, при этом образуется вода.
Баланс. Расчеты энергетического баланса показали, что при расщеплении глюкозы гликолитическим путем и через цикл Кребса с последующим окислением в дыхательной цепи до СО2 и Н2О на каждую молекулу глюкозы образуется 38 молекул АТФ. Причем максимальное количество АТФ образуется в дыхательной цепи – 34 молекулы, 2 молекулы - в ЭМП-пути и 2 молекулы – в ЦТК (рис. 36).
|

Неполное окисление органических соединений
Дыхание обычно связано с полным окислением органического субстрата, т.е. конечными продуктами распада являются СО2 и Н2О.
Однако некоторые бактерии и ряд грибов не до конца окисляют углеводы. Конечными продуктами неполного окисления являются органические кислоты: уксусная, лимонная, фумаровая, глюконовая и др., которые аккумулируются в среде. Этот окислительный процесс используется микроорганизмами для получения энергии. Однако общий выход энергии при этом значительно меньший, чем при полном окислении. Часть энергии окисляемого исходного субстрата сохраняется в образующихся органических кислотах.
Микроорганизмы, развивающиеся за счет энергии неполного окисления, используются в микробиологической промышленности для получения органических кислот и аминокислот.