

Paleo / MARTYNOV
.pdfразнообразия гидробионтов. Новые экологические гильдии почти мгновенно, в масштабах геологического времени, осуществили глобальную колонизацию дна и водной толщи морей, что сопровождалось формированием многочленистой трофической структуры экосистем. Среди беспозвоночных главную роль играли древние группы, но все большее значение приобретали животные с карбонатным скелетом: строматопоры, табуляты, гелиолитоидеи, ругозы, иглокожие (морские лилии, морские пузыри).
Трилобиты были менее разнообразны по числу родов, но сохранили одно из ведущих мест по численности. Малочленистые (Miomera) кембрийские формы редки. Среди многочленистых (Polymera) доминировали формы с прочным и толстым известковым панцирем; все они приобрели способность свертываться в шарик, защищая мягкую брюшную мембрану. Как следствие этого – развитие равновеликих и близких по очертаниям цефалона и пигидия. Ордовикские трилобиты обычно имеют небольшое и постоянное в пределах родственной группы число туловищных сегментов, обладают не только опистопарными, но очень часто пропарными лицевыми швами (Asaphus, Illaenus, Pterygometopus и др.) (Рис. 14: 3 – 6).
Характерными и массовыми в ордовике были граптолиты (вымерший класс Graptolithina).*
Рис. 10. Класс Graptolithina – Граптолиты. Образ жизни граптолитов: а, б – планктон
(подкласс Graptoloidea); в, г – бентос (подкласс Stereostolonata)
* Таксономический ранг граптолитов и их положение в системе животного мира дискусионны. Их включали в качестве самостоятельного класса к кишечнополостным, либо понижали ранг и относили к гидроидным полипам, существует также мнение о сходстве граптолитов и мшанок. Класс граптолитов разделяется на два подкласса – Стереостолонаты – Stereostolonata – представлены древовидными или сетчатыми колониями и Граптолоидеи – Graptoloidea – колонии прямые, изогнутые или спиральнозавитые с одним или двумя рядами тек.
Из современных форм граптолиты имеют сходство с некоторыми современными полухордовыми из класса крыложаберных – Pterobranchia (ранее объединяли с ними) – например представители рода Rhabdopleura.
41
Граптолиты были морскими организмами. Они прикреплялись к дну, либо обитали в толще воды. О планктонном образе жизни свидетельствуют находки воздушного пузыря – пневматофора. Некоторые колонии прикреплялись к плавающим объектам (псевдопланктон). Колонии граптолитов называются рабдосома, они были достаточно разнообразны по форме и размерам (Рис. 10). Широко представлены кустистые колонии стереостолонат (Stereostolonata). Настоящие граптолиты в ордовике быстро эволюционировали и имели широкие ареалы, что делает их важнейшими руководящими ископаемыми ордовика. На основании изучения граптолитов отложения ордовика расчленяются на 19 граптолитовых зон (зона - наиболее дробное подразделение в стратиграфии). Для раннего и среднего ордовика характерны, главным образом, безосные формы из одинаковых ветвей, собранных в пучок (Phyllograptus, Didymograptus), для среднего и позднего ордовика – осеносные двурядные граптолиты (Diplograptus, Climacograptus и др.).
Брахиоподы были представлены как беззамковыми формами с хитиновофосфатной раковиной (Obolus), так и многочисленными замковыми с известковыми раковинами. Среди последних важную роль играли ортиды,
пентамериды и строфомениды (Orthis, Strophomena, Porambonites).
Из моллюсков широко распространены в ордовике головоногие – наутилоидеи (Nautiloidea) с прямой или слабо изогнутой раковиной – эндоцера-
тоидеи (Endoceratoidea, Endoceras), ортоцератоидеи (Orthoceratoidea, Orthoceras) и другие группы, которые вели придонный, активный хищный образ жизни. В ордовике они испытывают расцвет, их раковины часто достигали в длину до 9 м – это были самые крупные из раковин, когда-либо встречавшихся у беспозвоночных животных. Полость раковины была разделена на камеры, заполнявшиеся газом, что позволяло им держаться на плаву. Некоторые из ортоцератид, возможно, поедали остатки погибших животных, но большинство было активными хищниками. Передвигались они ”реактивным способом”, как и современные головоногие, с силой выбрасывая воду из мантийной полости.
С ордовиком, особенно со средним, связано второе массовое появление разнообразных многоклеточных, имевших известковый скелет, это различные кишечнополостные, головоногие моллюски, брахиоподы, иглокожие. Важной группой являлись кишечнополостные, представленные в ордовике строматопороидеями (Stromatoporata, вымерший подкласс Hydrozoa), в начале среднего ордовика появляются первые ругозы (вымерший подкласс кораллов – Tetracoralla, Rugosa), представленные простыми однозонными формами; появились гелиолиты (вымерший надотряд Heliolitoidea); среди табулят (Tabulatomorpha – вымерший подкласс кораллов, давший начало ругозам) развиты формы с несообщающимися между собой кораллитами (инкоммуникатные). Этот достаточно богатый и разнообразный мир бентосных кишечнополостных принимал активное участие в построении рифов (Рис. 11), сменив кембрийские археоциатово-водорослевые сообщества.
42

Рис. 11. Кишечнополостные рифообразователи ордовика. Тип Coelenterata, или Cnidaria – Кишечнополостные, или Стрекающие: Класс Hydrozoa – Гидроидные, подкласс Stromatoporata – Строматопораты, 4 – 7 внешняя форма колоний. 4 – полусферическая, 5 – сферическая, 6 – грушевидная, 7 – желваковидная с пальцевидными выростами, 8 – Lebechia (средний ордовик – девон) – поперечный срез; 9 – Stromatopora (силур – мел) – поперечный срез; 10 – Actinostroma (девон). Класс Anthozoa – Коралловые полипы, подкласс
Tabulatomorpha – Табулятоморфы. 1 – 3 надотряд Chaetetoidea (ордовик – неоген) 1 –
внешний вид колонии с плоскостями перерыва роста, 2 – поперечный срез, 3 – продольный срез; 11 – 20 надотряд Tabulatoidea (средний кембрий – пермь), 11 – 13 отряд Favositida (средний ордовик – пермь) 11 – внешний вид колонии фавозитид, 12 – поперечный срез Favosites gothlandicus (с – поры), 13 – продольный срез; 18 – 20 отряд Syringoporida (средний ордовик – ранняя пермь) 18 – Syringopora – общий вид колонии, 19 – 20 поперечный и продольный срезы; 14 – 15 надотряд Heliolitoidea (средний ордовик – средний девон) Heliolites s. lato – поперечный и продольный срез. Подкласс Tetracoralla, или Rugosa – Четырехлучевые кораллы, или Ругозы 16 – 17 – Favistina (средний – поздний ордовик) поперечный и продольный срез однозонного коралла
В ордовике наблюдается настоящий расцвет иглокожих, в этот период они представлены почти 20 классами, в том числе и всеми современными. Иглокожие принимали активное участие в донных биоценозах. Это были преимущественно морские пузыри (класс Cystoidea), образующие обширные
43

поселения, и в меньшей степени морские лилии (класс Crinoidea), морские звезды (Asterozoa). Морские пузыри – исключительно палеозойская группа древних иглокожих. Их мешковидное или шаровидное тело прикреплялось к субстрату при помощи зачаточного стебелька или свободно лежало на дне (Рис. 12). Ротовое отверстие находилось в центре и у некоторых видов было окружено венчиком коротких, членистых брахиолей. Цистоидеи дали начало морским бутонам (Blastoidea) и морским лилиям (Crinoidea).
Рис. 12. Тип Echinodermata – Иглокожие, класс Cystoidea – Морские пузыри: 1 - Echinosphaerites (средний – поздний ордовик) а – анальная пирамидка, р – ротовое отверстие, с - стебелек; 2, 3 - Heliocrinites balticus (средний ордовик) а – анальная пирамидка; 4 – Proteroblastus (силур) р – рот, ab – амбулакральная борозда с брахиолями, с – место прикрепления к субстрату; 5 – Glyptosphaerites leuchtenbergi (средний ордовик) а – пищевые желобки, b – следы прикрепления брахиолей, р – ротовое отверстие, прикрытое пятью пластинками, ао – анальное отверстие; 6 – Aristocystis (силур) а – анальный конус, р – половая пора, с – место прикрепления к субстрату; 7 – Echinoencrinites senckenbergii (ордовик) а – анальное отверстие, р – поровые ромбы, с - стебель
Значительно увеличивается численность фораминифер и радиолярий, которые становятся породообразователями. В пресных и солоноватоводных водоемах появляются гигантские хищные хелицеровые (до 2 – 3 м длиной) Eurypterida. Они вели плавающий придонный и ползающий донный образ жизни. В пресных водоемах появляются первые Oligochaeta.
Помимо этих основных групп беспозвоночных, в морях ордовика существовали и другие, которые не пользовались столь широким распространением, различные губки и черви (остатки которых известны в виде челюстных образований – сколекодонтов), остракоды, двустворки и гастроподы,
44
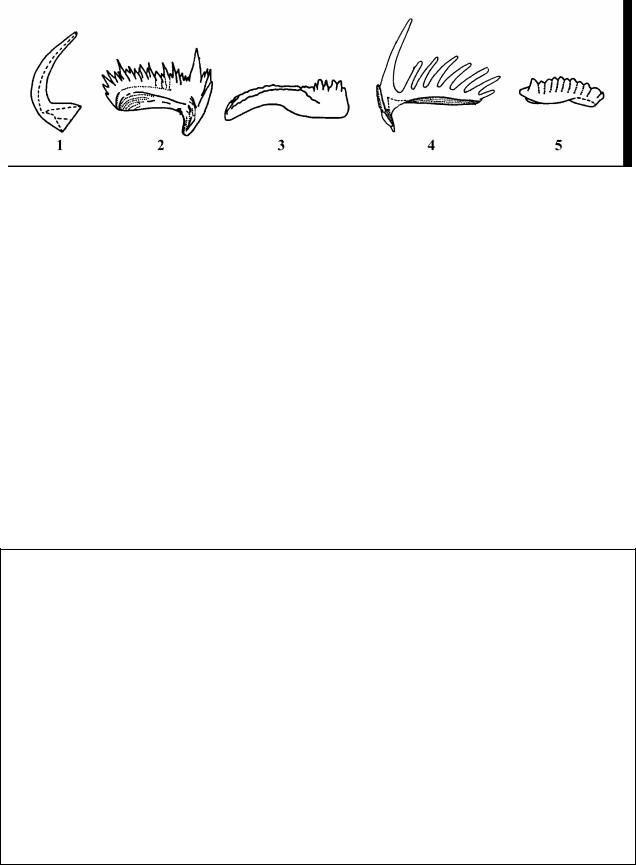
морские и пресноводные мшанки (строившие рифы вместе с кораллами), загадочные зубоподобные образования – конодонты (Рис. 13).*
Рис. 13. Неопределенный тип ? Chordata – Хордовые, класс Conodonti – Конодонты. Типы конодонтов: 1 – простой конический; 2, 4 – стержневидные; 3, 5 - платформенные
Позвоночные в ордовике хотя и немногочисленны, но уже представлены двумя подклассами бесчелюстных – Agnatha (телодонты Thelodonti и разнощитковые Heterostraci). Это были бесчелюстные рыбообразные животные длиной от 2 см до 2 м. Они были лишены челюстей, имели парное носовое отверстие, непарные плавники (из парных плавников иногда присутствовали грудные), внутренний скелет был хрящевым. Туловище бесчелюстных было покрыто шагренью из кожных щитков и чешуевидных пластин, которые могли срастаться, образуя панцирь, покрывающий голову и переднюю часть туловища; задняя часть туловища и хвост были покрыты чешуей. Телодонты и большинство разнощитковых вели придонный малоподвижный образ жизни. Небольшая часть разнощитковых с веретенообразным телом небольших размеров могла вести нектонный образ жизни. Их потомки, миноги и миксины, живущие в настоящее время, утратили наружный скелет и перешли к паразитическому образу жизни.
* В 1856 г русский палеонтолог Христиан Пандер описал ископаемые микроскопические зубовидные образования, которые он назвал конодонты. С тех пор систематическое положение вымерших морских организмов, которым принадлежали конодонты, является предметом дискуссии. В ископаемом состоянии их остатки представлены в основном разнообразными по форме зубоподобными элементами размером от долей мм до 1 мм, редко 3 – 4 мм (Рис. 1). По составу это фосфат кальция. Находки отпечатков организмов конодонтоносителей единичны. Особенности образа жизни неизвестны. Предполагают, что конодонтоносители были близки к щетинкочелюстным (Chaetognatha), которые широко распространены в современных бассейнах. Конодонты встречаются в морских отложениях от среднего кембрия до триаса. В настоящее время рассматриваются как отдельный подтип Conodontochordata – Конодонтохордаты, или класс Conodonti – Конодонты в пределах Хордовых. Систематика конодонтов полностью искусственна. Большинство таксонов установлено на основе морфологии изолированных элементов, реже аппаратов. Всего описано около 200 родов. Вследствие быстрых темпов эволюции конодонтовых аппаратов и широкого распространения в палеозойских морях они имеют важное значение для определения возраста осадочных пород. Окраску конодонтовых элементов используют для определения палеотемператур.
45
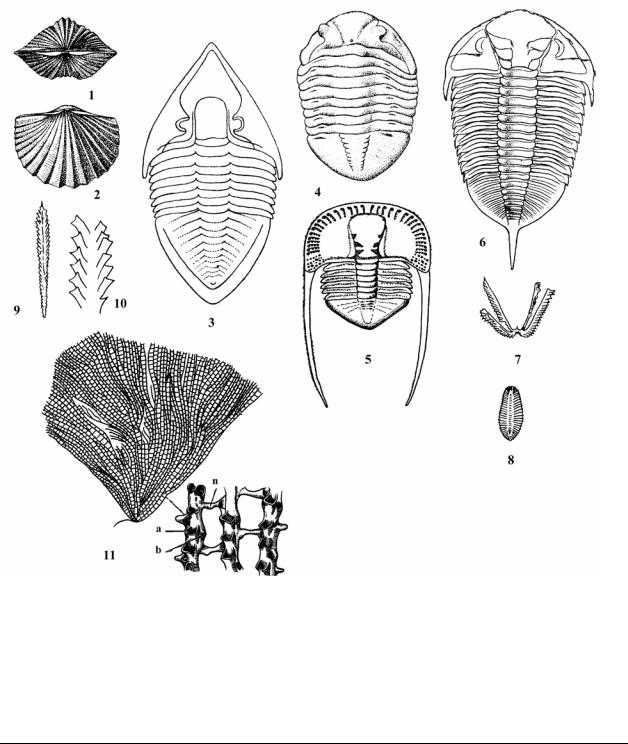
Рис. 14. Характерные ископаемые беспозвоночных ордовика: тип Brachiopoda – Брахиоподы. 1 – 2 – Platystrophia lynx (средний ордовик). Подтип Trilobitomorpha – Трилобиты, п/к Polymera – Многочленистые; 3 – Megistaspis limbata (ранний ордовик); 4 – Asaphus expansus (ранний ордовик); 5 – Cryptolithus ornatus (средний ордовик); 6 – Dilmaritina socialis (средний и поздний ордовик). Класс Graptolithina – Граптолиты, п/к Graptoloidea –
Граптолоидеи, 7 – Tetragraptus serra (ранний ордовик); 8 – Phyllograptus densus (ранний ордовик); 9-10 – Orthograptus amplexicaulis (средний и поздний ордовик). п/к Stereostolonata – Стереостолонаты, 11 – Dictyonema coalitum (ранний ордовик), a – автотеки, b – битеки, n - перемычки
В конце ордовика наблюдается первое массовое вымирание многих древних групп беспозвоночных, в результате которого исчезло около 35% семейств и более 50% родов морских животных. Но в отличие от более поздних “великих вымираний” оно не сопровождалось полным вымиранием каких-либо экологических гильдий. В раннем силуре позднеордовикское снижение таксономического разнообразия быстро компенсировалось быстрым восстановлением новых экологических гильдий. Из ордовика известны остатки низших растений – зеленых водорослей и грибов; из высших – мхи и, возможно, плауновидные. Судя по находкам своеобразных спор, в карадокском веке на суше уже существовали наземные растения.
46
СИЛУРИЙСКИЙ ПЕРИОД (СИСТЕМА)
Силурийская система была установлена в 1835 г английским ученым Р. Мурчисоном на п-ове Уэльс в Великобритании и названа по имени силуров, древнего племени, населявшего эту территорию. Силур – самый непродолжительный период палеозоя (35 млн. лет), начался 435 и закончился 400 млн. лет назад. В пределах силура принято выделять два отдела (Табл. 5).
Материковые платформы Лаврентия и Балтика в этот период соединились в новый блок – Еврамерику, отделенный от Гондваны небольшим океаном Рея. В области их соединения возникли горные системы: Аппалачский хребет и горы Скандинавии. С севера к Еврамерике приблизились Сибирь и Казахстан. Следствием усиления горообразовательных процессов (продолжался период каледонской складчатости) во второй половине силура стало обширное осушение материковых платформ (регрессия моря). Климат силура был более аридный чем ордовика. В раннем силуре предполагается существование двух биогеографических областей: одной на северных материках и в Австралии, другой – в Южной Америке и Африке. Первая характеризуется остатками богатой и разнообразной теплолюбивой фауны, вторая – своеобразным обедненным комплексом.
Таблица 5
Геохронологическое (стратиграфическое) подразделение силурийской системы
Отдел |
Ярус |
Верхний |
Пржидольский S2 p |
|
Лудловский S2 ld |
Нижний |
Венлокский S1 w |
|
Лландоверийский S1 l |
ОРГАНИЧЕСКИЙ МИР
Ордовикский биотический кризиз обусловил обедненный характер фауны раннего силура. В первую очередь это сказалось на трилобитах, число которых резко сократилось. Лишь во второй половине периода разнообразие морской биоты несколько возросло. В целом органический мир силура стал богаче и разнообразнее, чем в ордовике. Одним из важнейших событий этого периода было появление наземных растений. Освоение растениями наземных пространств явилось важным этапом развития органического мира. На суше обитали грибы, мхи и высшие растения – риниофиты, а в конце силура появились примитивные плауновые. Кроме растений на суше встречались многоножки и скорпионы. В целом органический мир силура, как и ордовика, характеризуется доминированием древних групп – граптолитов, трилобитов, строматопорат, табулят, древних семейств брахиопод, головоногих моллюсков с прямой раковиной.
47
Органический мир моря. В морях силурийского периода главенствующее положение занимали те же группы беспозвоночных животных, которые доминировали и в ордовике, но появились их новые представители (Рис. 15). Среди полухордовых по-прежнему очень широко распространены граптолиты. На смену безосным граптолитам, вымершим в конце ордовика, в раннем силуре приходят осеносные граптолиты в начале с двурядными, а позднее с однорядными колониями. В конце раннего силура исчезают двурядные и сетчатые граптолиты и в позднем силуре господствующее положение занимают граптолиты с однорядными колониями. В конце силура или начале девона собственно граптолиты вымирают. Кишечнополостные – рифостроители представлены строматопороидеями, несообщающимися и сообщающимися табулятами (Halysites, Paleofavosites и др.), гелиолитами. Появились двузонные четырехлучевые кораллы; в отличие от однозонных, у них развита пузырчатая ткань, часто заполняющая всю внутренность теки (Goniophyllum и др.). Головоногие моллюски представлены группами, известными и в ордовике; правда, они (особенно эндоцератоидеи) играли менее заметную роль. Появились первые бактритиды (подкласс Bactritoidea), ключевая группа в эволюции головоногих. Это небольшая группа головоногих моллюсков с прямой или слегка изогнутой раковиной просуществовала с силура до триаса и дала начало аммонитам (п./к. Ammonoidea) и внутреннераковинным (п./к. Endocochlia). Предполагают, что бактриты произошли от наутилоидей с прямой раковиной (возможно актиноцератоидей).
Среди брахиопод наряду с отрядами, типичными для ордовика, появились новые, широко известны атрипиды (Atrypida), спирифериды (Spiriferida), ринхонеллиды (Rhynchonellida), появились первые продуктиды
(Productida – Atrypa, Conchidium, Eospirifer и др.). Таким образом, для силу-
ра характерно появление брахиопод со сложными поддержками ручного аппарата. Многочисленные брахиоподы часто выступали в роли породообразователей. Иглокожие были представлены в подавляющем большинстве прикрепленными формами, но цистоидей становилось меньше, зато увеличилось значение морских лилий (Crinoidea), появились бластоидеи (класс Blastoidea –морские бутоны). Трилобиты встречаются в силурийских породах в гораздо меньшем количестве. Конодонты стали разнообразнее. Среди фораминифер появляются первые представители с известковой раковиной (ранее существовали только агглютинированные формы).
Помимо основных групп, в морских отложениях силура обычны губки, черви, остракоды, эвриптериды (Eurypterida – водные скорпионы), двустворки и гастроподы, древнейшие тентакулиты, мшанки, морские ежи и звезды. Со второй половины силура позвоночные организмы (бесчелюстные) начали осваивать континентальные пресноводные бассейны. В позднем силуре появляются первые настоящие рыбы, снабженные челюстями, парными плавниками и более совершенным внутренним скелетом.
48
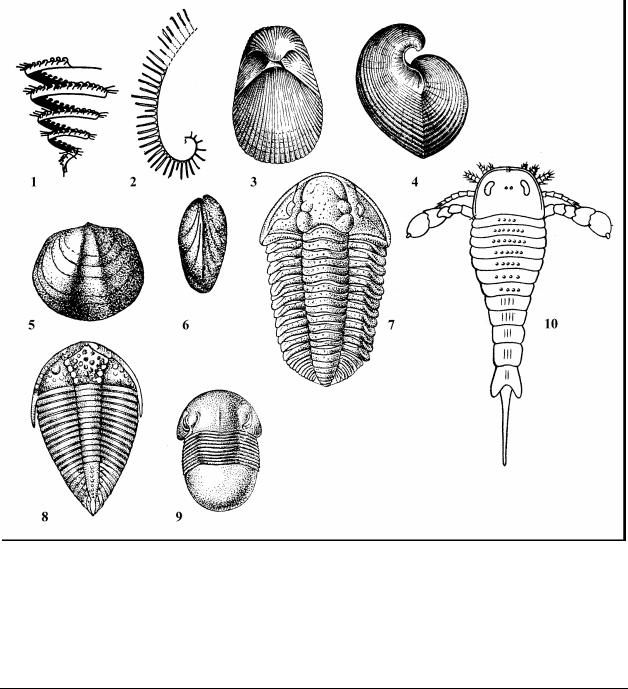
Рис. 15. Характерные ископаемые беспозвоночные силурийского периода: Класс Graptolithina – Граптолиты, п/к Graptoloidea – Граптолоидеи, 1 – Spirograptus (ранний силур); 2
– Rastrites (ранний силур). Тип Brachiopoda – Брахиоподы, 3-4 – Conchidium knighti (позд-
ний силур); 5-6 – Stricklandia lens (ранний силур). Подтип Trilobitomorpha – Трилобиты,
п/к Polymera – Многочленистые, 7 – Calymene blumenbachii; 8 – Encrinurus punctatus; 9 –
Bumastus barriensis. Подтип Chelicerata – Хелицеровые, п/к Merostomata, 10 – Eurypterus fischeri (поздний силур – ранний девон)
Они были представлены акантодами (колючезубыми, класс Acanthodei) - рыбами, соединявшими в себе признаки хрящевых и костных рыб. Это были первые на Земле хищные рыбы. Тело акантод было покрыто плотными квадратными чешуйками, сходными с ганоидными. Акандоды имели один анальный и один или два спинных плавника, между парными грудными и брюшными плавниками имелась серия дополнительных плавников, снабженных впереди колючками - ихтиодорулитами, внутренний скелет был хрящевым. У ранних форм число парных плавников на нижней части туловища могло доходить до семи, что служит доказательством происхождения парных плавников из непрерывных кожных складок. Акантоды появились в позднем силуре, были распространены в девоне и вымерли в начале перми.
В силуре продолжают развиваться бактерии, грибы, водоросли – синезе-
49
леные, багряные и зеленые.
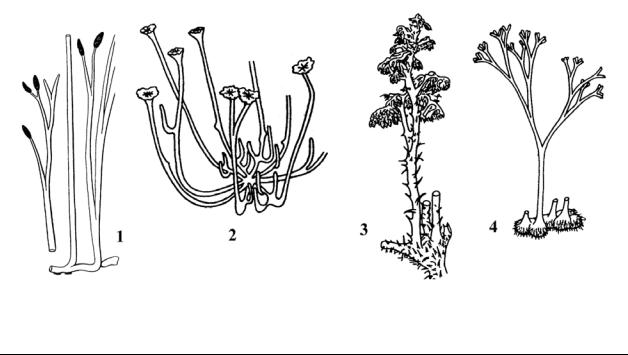
Органический мир суши. Конец силура и начало девона ознаменовались крупными горообразовательными процессами, которые привели к увеличению площади суши, созданию новых горных цепей и регрессии моря. В конце силура появляются первые наземные растения - риниофиты и примитивные плауновидные (Barragvanathia). Риниофиты, или Риниевые (Отдел Rhyniophyta) – своеобразные споровые растения, напоминающие современные плауны. Риниофиты были лишены корней, а в основном и листьев (Рис. 16). Они состояли из ветвившихся зеленых стеблей высотой до 30 см и горизонтально тянувшегося в почве корневища. Риниевые, как первые достоверные растения суши, создавали целые зеленые ковры на влажной почве по берегам водоемов. В связи с этим первую наземную флору часто называют псилофитовой или проптеридофитовой. Наземная фауна пополнилась скорпионами (Palaeophonus – по мнению некоторых авторов он обитал в водной среде).
Рис. 16. Первые высшие наземные растения. Отдел Rhyniophyta – Риниофиты, или Риниевые (силур – девон): 1 – Rhynia (ранний девон) внешний вид спорофита; 2 – Sciadophyton (ранний девон) внешний вид гаметофита; 3 – Psilophyton (ранний – средний девон); 4 – Horneophyton (ранний девон) реконструкция
Выход растений на сушу и завоевание ими в девоне огромных наземных пространств явился важнейшим этапом развития органического мира, открывшим широкие возможности для прогрессивной эволюции. Начался новый этап эволюции биосферы Земли, количество живого вещества перестало определяться площадью поверхности океанов, что привело к значительному увеличению биомассы всей Земли. Как известно, современная биомасса организмов суши едва ли не в 800 раз превышает биомассу Мирового океана. Сухопутные животные составляют 93% современного видового состава, водные только 7%. Аналогичные значения мы видим и среди растений – 92% видов наземные и лишь 8% водные.
50