

Paleo / MARTYNOV
.pdfПо ископаемым сообществам растений установлено, что смена палеозойской флоры мезозойской проходила на земной поверхности не одновременно. На территории современной Европы существенное изменение флоры произошло на рубеже ранней и поздней перми (именно в это время здесь наступила резкая аридизация климата), на территории современной Сибири - на рубеже перми и триаса и в Гондване - на рубеже раннего и среднего триаса. Климат в области распространения вестфальской флоры становится более сухим, вызывая затухание процессов углеобразования и распространение ксерофитной флоры. В тунгусской и гондванской областях в пермское время климатические условия изменяются сравнительно мало, поэтому здесь пермские флоры обладают многими чертами сходства с позднекаменноугольными; во многих местах продолжаются процессы углеобразования (Северный Урал, Кузбасс, Китай, Индия, Австралия). К концу пермского периода в эти области проникают европейские формы. Флора приобретает новый облик, в котором господствующее положение принадлежит папоротникам и голосеменным.
Изменения в наземной фауне протекали в целом не столь драматично, хотя и здесь происходят существенные изменения. Аридизация климата, появление на больших территориях ландшафтов типа современных степей и пустынь, в первую очередь отразилась на изменении состава растительности и позвоночных. Если в начале перми были еще обильны и разнообразны земноводные (стегоцефалы), то к концу периода многие из них вымерли, часть из них с запозданием пытается закрепиться на суше. Создались благоприятные условия для развития пресмыкающихся, наиболее приспособленных в то время к наземному образу жизни. В пермский период начинается расцвет пресмыкающиеся, они достигают большого разнообразия в своих приспособлениях к окружающей среде. Среди них известны растительноядные, хищные, всеядные и, по-видимому, моллюскоядные представители, которые быстро осваивали новые местообитания, вытесняя хуже приспособленных конкурентов из числа стегоцефалов и батрахозавров. Увеличивается разнообразие парарептилий (класс Parareptilia - котилозавры, сеймуриаморфы) и особенно рептилий (синаптозавры, лепидозавры, текодонты, зверообразные) (Рис. 28).
В конце перми происходит одно из наиболее катастрофических вымираний морских организмов, затронувшее 95% видового состава и около половины семейств беспозвоночных. Но как ни странно, среди наземных и водных позвоночных ничего сравнимого по масштабам в это время не наблюдалось. По своим объемам пермское вымирание принадлежит к категории так называемых "великих вымираний" и сравнимо с широко известным великим вымиранием в конце мелового периода. Причины вымираний, приводившие к радикальным изменениям общего облика фауны и флоры, остаются еще загадочными и на протяжении десятков лет не перестают быть объектами дискуссий и соревнования гипотез, пытающихся объяснить эти события. Вымирание вообще сопровождает эволюцию организмов, как ее неизбежный побочный результат. При этом следует отметить и тот факт, что под
71
вымиранием в палеонтологии подразумевают два совершенно разных процесса: прекращение существования таксона, не оставляющего потомства, и его превращение путем филетической эволюции в другой таксон, отличающийся определенными признаками.
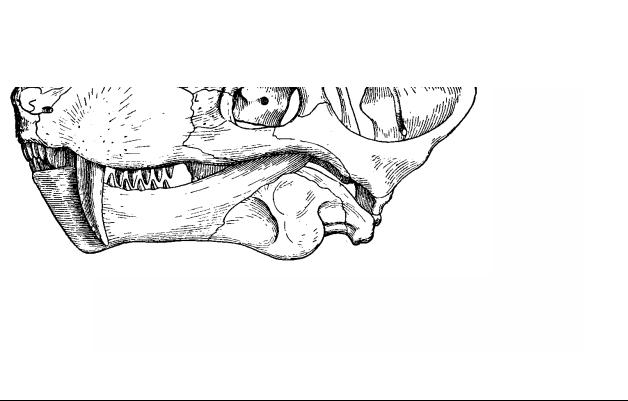
Рис. 28. Подкласс Theromorpha, или Synapsida – Зверообразные, или Синапсиды. Череп зауроктона (верхняя пермь) – Sauroctonus progressus (обратите внимание на дифференцированность зубной системы)
Последний процесс называют ложным вымиранием. Вымирание происходило на любом отрезке эволюционной истории органического мира, либо в результате конкуренции с биологически близкими видами, либо по причине неспособности вида выдержать те или иные изменения абиотических факторов внешней среды и т. д. (Табл. 9). В принципе эти же причины определяют фатальную судьбу каждого из вымирающих видов и в эпохи великих вымираний, но тогда одновременно обрывается множество филетических линий. Следует учитывать тот фактор, что вымирание может происходить достаточно постепенно, но в огромной исторической перспективе геологического времени такая постепенность нередко выглядит относительно быстрым и резким изменением фауны и флоры. Например, по некоторым данным только за последние триста лет исчезло свыше 150 видов позвоночных, безусловно, ведущую роль в этом играет человек, но при таких темпах страшно представить масштабы, протекающего в настоящее время вымирания в геологически заметные сроки (хотя бы несколько миллионов лет).
О причинах пермского вымирания было выдвинуто несколько гипотез, большинство из которых в конечном итоге опирается на теорию континентального дрейфа. Согласно гипотезе, предложенной Д. Валентайном и Э. Мурсом (1974), образование Пангеи обусловило сокращение общей площади мелководных бассейнов в области континентального шельфа, которая является наиболее благоприятной средой для жизни многих групп морских организмов, это должно было привести к существенному нарушению многих сложившихся в средине палеозоя морских биоценозов, и как следствие, к вымиранию многочисленных видов.
Другая гипотеза, предложенная Р. Боуэном (1975), обращает внимание на
72

потепление климата и завершение великого оледенения Гондваны во второй половине перми, что могло быть связано с относительно небольшим смещением входивших в ее состав континентальных плит, в результате которого Южный полюс оказался в океане. Потепление климата привело к бурному таянию ледников и поступлению в океаны огромного объема талых вод, что изменило соленость морской воды. С. Стевенс (1977) подчеркивал тот факт, что пермское вымирание отразилось в первую очередь именно на тех группах морских организмов, которые наиболее чувствительны к изменению солености воды: кораллы, морские лилии, мшанки, брахиоподы, головоногие моллюски и др. Новая стабилизация температурного и солевого режимов в океане произошла уже в триасовом периоде, и разнообразие форм морских беспозвоночных к концу триаса вновь стало возрастать.
Таблица 9
Основные пики вымирания в морских сообществах
Наблюдаемый пик вымирания, млн.. лет назад
Третичный период |
Средний миоцен |
11,3 |
|
Поздний эоцен |
38 |
Меловой период |
Маастрихт |
65 |
|
Сеноман |
91 |
|
Готерив |
125 |
Юрский период |
Титон |
144 |
|
Келловей |
163 |
|
Байос |
175 |
|
Плинсбах |
194 |
Триасовый период |
Норий |
219 |
|
Оленек |
243 |
Пермский период |
Джульфа |
248 |
Несколько иначе, подходят к решению этого вопроса, сторонники вулканической теории. По их мнению, причиной кризиса была активная вулканическая деятельность. Извержение вулканов сопровождалось выбросами в атмосферу диоксида серы и оксида азота, послуживших причиной интенсивных кислотных дождей. У большинства палеозойских организмов репродуктивный цикл происходил в водной среде с нейтральным уровнем рН. В водоемах с кислой средой гаметы и молодь организмов погибали.
73

ИСТОРИЯ ЗЕМЛИ В МЕЗОЗОЙСКУЮ ЭРУ (ЭРАТЕМА, ГРУППА)
(греч. mesos – средний, промежуточный; zoe - жизнь)
ОБЩИЕ СВЕДЕНИЯ Мезозойская эра получила свое название благодаря ее промежуточному
положению между древней – палеозойской и новой – кайнозойской эрами. Начало около 250 млн. лет, конец – 65 млн. лет назад, продолжительность эры около 185 млн. лет. Мезозойская эра подразделяется на три периода: триасовый, юрский и меловой. Отложения этих периодов составляют соответствующие системы, которые образуют мезозойскую группу.
В южном полушарии в мезозое распался древний сверхконтинент Пангея, включавший в себя почти всю сушу Земли. Начался процесс формирования современных очертаний материков и океанов. Возник океан Тетис, разделивший материки Лавразию и Гондвану. Изоляция материков явилась предпосылкой расхождения биот.
ОРГАНИЧЕСКИЙ МИР
Органический мир мезозоя очень своеобразен, он занимает промежуточное положение между палеозойским и кайнозойским. Это было время смены старых, более примитивных жизненных форм новыми, более прогрессивными. Для этой эры характерен новый расцвет большинства типов животных и отделов растений, появившихся ранее.
В морях широчайшее развитие получили аммоноидеи нового систематического состава и белемниты (особенно в юре и мелу). Резко увеличилось разнообразие двустворок и гастропод, брахиоподы стали менее часты, появились шестилучевые кораллы. Появляются новые морские ежи, морские лилии, мшанки, фораминиферы также обновили состав. Морские позвоночные претерпели коренные изменения: изменился состав рыб, появились и распространились крупные плавающие рептилии – ихтиозавры, плезиозавры, мозазавры.
Особым своеобразием отличался органический мир суши, где доминировали различные пресмыкающиеся, в особенности динозавры,* среди которых было много гигантских форм. Никогда рептилии не играли такой роли в наземных и морских ценозах как в мезозое. В связи с чем, мезозой по праву называют эрой рептилий. Появившиеся в конце триаса млекопитающие до конца мезозоя оставались мелкими животными, не игравшими существенной роли в наземных ценозах. В триасе появились первые птицы.
*Название “динозавры” (страшные ящеры) относится к нескольким независимым группам рептилий из подкласса архозавров (Archosauria) и в современном понимании не имеет определенного таксономического статуса. До настоящего времени среди палеонтологов нет единого мнения в отношении классификации динозавров. Наиболее распространено распределение этих животных по двум отрядам: ящеротазовых (Saurischia) и птицетазовых (Ornithischia), но современный материал не всегда позволяет провести такое разграничение.
74
Наземный растительный мир отличался расцветом голосеменных и папоротникообразных, в мелу к ним присоединяются покрытосеменные – цветковые. Очередной расцвет растений, среди которых было много древесных форм, привел к новой эпохе углеобразования. Особенно активно процессы углеобразования проходили в юре. Органический мир мезозоя обнаруживает поясную климатическую зональность, особенно в поздней юре, что затрудняет корреляцию стратиграфических шкал разных поясов.
ТРИАСОВЫЙ ПЕРИОД (СИСТЕМА)
Был выделен в 1831 г под названием "кейперских отложений" бельгийским ученым Ж. д' Омалиусом д' Аллуа на территории Западной Европы, в Германской впадине. В 1834 г немецкий геолог Ф. Альберти назвал эти отложения триасом (в отличие от пермской системы, которую тогда в Западной Европе называли диасом). Триасовый период начался около 250 млн. лет назад и закончился 215 млн. лет назад, общая продолжительность периода 35 млн. лет. Разработка ярусной шкалы триаса производилась в Альпах, где были установлены все ныне существующие ярусы среднего и верхнего отделов, так как в Германском бассейне широко распространены континентальные отложения и выделенные там подразделения имеют исключительно местное значение (Табл. 10). Нижний отдел триаса не имеет общепринятого ярусного деления. На территории бывшего СССР его делят на два яруса: индский и оленекский.
Таблица 10
Геохронологическое (стратиграфическое) подразделение триаса
Эпоха (Отдел) |
Век (Ярус) |
Верхний триас |
Рэтский T3 r |
|
Норийский T3 n |
|
Карнийский T3 k |
Средний триас |
Ладинский T2 l |
|
Анизийский T2 a |
Нижний триас |
Оленекский T1 o |
|
Индский T1 i |
Суша в триасе представляла собой огромный континент Пангею, протянувшийся от Южного до Северного полюса. В целом триасовый период характеризуется морской регрессией. Только в позднем триасе началась трансгрессия. Климатическая обстановка раннего триаса изменилась по сравнению с пермским периодом незначительно. Наблюдается дальнейшее расширение аридных зон, к концу периода в отдельных районах климат стал влажным (гумидным), что привело к угленакоплению в Западной Сибири (Челябинский бассейн), Индокитае, Южном Китае и т. д. На значительных площадях сохранились аридные условия. Постепенно исчезла резко выраженная в перми широтная климатическая зональность.
75

ОРГАНИЧЕСКИЙ МИР
Триасовый период, первый период мезозойской эры, характеризуется значительным обновлением состава фауны и флоры за счет появления новых групп. В то же время в составе органического мира триаса сохраняются и типично палеозойские группы; среди беспозвоночных – брахиоподы (спирифериды), наутилоидеи с прямой раковиной, среди позвоночных – стегоцефалы, среди растений – каламитовые и др. Однако элементы нового, типичного для мезозоя, явно преобладали в триасе. В целом изменения органического мира на рубеже перми и триаса (т.е. между палеозоем и мезозоем) не носили катастрофического характера, были длительными, постепенными, но весьма существенными.
Органический мир моря. Из беспозвоночных были широко распространены аммоноидеи (Ammonoidea) – цератиты (Ceratitida), испытавшие несколько эволюционных подъемов и спадов. На рубеже перми и триаса цератиты представлены всего двумя родами, в оленекском веке их число достигает 113, в ладинском резко падает до 50, в карнийском вновь возрастает до
140.
Рис. 29. Подкласс Ammonoidea – Аммоноидеи, отряд Ceratitida (пермь – триас): а – в – Paraceltites (ранняя пермь); г – е – Ceratites (средний триас), зл – зазубренная лопасть, V – брюшная лопасть
Цератиты в триасе занимают главенствующее место среди морских беспозвоночных как по числу родов (примерно 450), так и по форме и характеру скульптуры раковины, но ни один из многочисленных родов не перешел в юру (Рис. 29). В норийском веке (верхний триас) цератиты полностью вымирают, и им на смену приходят первые настоящие аммониты с аммонитовой лопастной линией.
76

Рис. 30. Руководящие ископаемые триасового периода: 1 – Meekoceras boreale (ниж-
ний триас); 2 – Halobia neumayri (верхний триас); 3 – Doricranites bogdoanus (ранний триас); 4 – Ceratites nodosus (средний триас); 5, 6 – Monotis ochotica (верхний триас); 7 – Myalina dalailamae (ранний триас); 8, 9 – Eumorphotis ivanovi (ранний триас)
В меньшей степени были распространены другие цефалоподы (Cephalopoda): наутилиды (Nautiloidea), ортоцератиды (Orthoceratoidea), белемниты
(Belemnitida). Наутилиды были представлены теми же подотрядами, что и в перми, но возникли новые роды. Появившиеся еще в позднем карбоне белемниты (хотя проблематичные формы известны еще из девона) продолжают свое развитие, но в триасе были очень редки и принадлежали к отряду Aulacocerida.
77
Сильно изменяется в триасе фауна двустворчатых и брюхоногих моллюсков, увеличилось их многообразие, двустворки занимают экологические ниши, которые в палеозое были заняты брахиоподами. Среди двустворок появился подотряд Ostreina (устрицы), много новых семейств. Многие виды родов Halobia, Daonella, Monotis, Claraia являются руководящими ископаемыми (Рис. 30). Среди брюхоногих начался расцвет отряда Mesogastropoda.
В конце перми вымерли четырехлучевые кораллы, а со среднего триаса им на смену приходят склерактинии (Scleractinia) – первые представители подкласса шестилучевых кораллов (Hexacoralla). И хотя в триасе шестилучевые кораллы еще не получили большого разнообразия, но в дальнейшем широко распространились в юрских и меловых морях, став одним из основных рифообразователей.
Из палеозойских брахиопод в триасе в значительном количестве сохранились спирифериды, но доминировали теребратулиды (Terebratulida) и ринхонеллиды (Rhynchonellida). Большинство палеозойских отрядов мшанок вымерло в конце перми, некоторые доживали в триасе. Существовавший в палеозое отряд Cyclostomata в триасе стал более разнообразным по составу.
Иглокожие также испытали перемены. Лишь некоторые архаичные морские лилии дожили до конца триаса. В начале периода появился новый подкласс Articulata, существующий и ныне. Древние морские ежи вымерли в конце палеозоя, сохранились только представители отряда Cidaroida – “копьеносные” ежи, передвигавшиеся на длинных иголках, достигшие в триасе и особенно позже большого многообразия. Появились диадемовые ежи (Diadematoida – правильные морские ежи), передвигавшиеся при помощи амбулакральных ножек.
Сильно изменился состав фораминифер; их число значительно сократилось, кроме представителей нодозариид (Nodosaria), которые стали доминировать среди фораминифер.
Морские позвоночные триаса более высоко развиты, чем их предшественники. Лучеперые рыбы (Osteichthyes, Actinopterygii) занимали промежуточное положение между хрящекостными и цельнокостными; костистые рыбы появились в среднем триасе. Триас ознаменовался успешным освоением рептилиями не только пресных водоемов, но и морей. Крупнейшими морскими хищниками были появившиеся в раннем триасе ихтиозавры (Ichthyosauria), в среднем – плезиозавры (Plesiosauria). Предполагают, что плезиозавры еще сохраняли связь с сушей, выползая на прибрежные отмели для откладки яиц, как это делают современные черепахи. Однако даже такое кратковременное пребывание на суше было достаточно затруднительно из-за огромных размеров (12 – 16 м) (Рис. 31). Ихтиозавры, по всей вероятности, уже не могли выходить на сушу и стали живородящими (Рис. 32). О живорождении ихтиозавров свидетельствуют и найденные скелеты крупных особей (самок?), внутри которых находились скелеты маленьких ихтиозавров, иногда свернутые кольцом (как зародыши в яйце), а также скелеты самок погибших в момент родов, скелет детеныша при этом выступал из клоаки хвостовым концом. Исходя из этого можно предположить, что у ихтиозавров
78
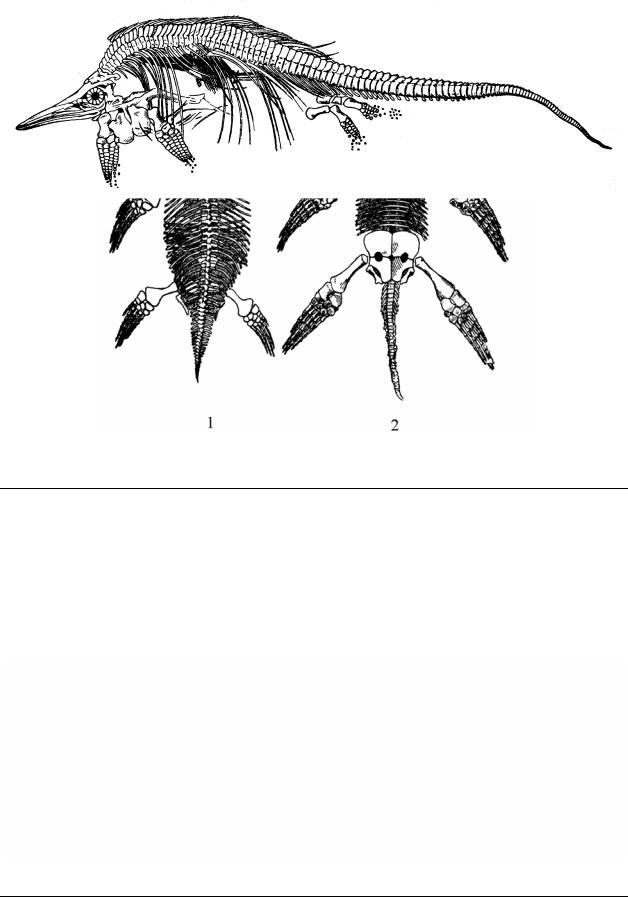
при родах, как и у современных китообразных, детеныши выходили хвостом вперед и сразу рефлекторно начинали работать хвостом.
Рис. 31. Подкласс Synaptosauria, или Euryapsida – Синаптозавры, или Эврипсиды. Скелеты плезиозавров: 1 – Cryptocleidus oxoniensis (верхняя юра) – вид сверху; 2 –
Thaumatosaurus victor (нижняя юра) – вид снизу
Происхождение специализированных водных рептилий – завроптеригий и плакодонтов, объединяемых в подкласс синаптозавров (Synaptosauria), так же как и ихтиозавров, до сих пор остается дискуссионным. Большинство палеонтологов склоняется к мысли об их независимом происхождении от ка- кой-либо группы котилозавров. Высказывалось также предположение о родстве ихтиозавров с черепахами. С позднего триаса известны древнейшие крокодилы и черепахи.
Рис. 32. Подкласс Ichthyosauria – Ихтиозавры, Стеноптеригиус – Stenopterygius quadriscissus (нижняя юра). Скелет самки со скелетом детеныша внутри брюшной части
79
Органический мир суши. В начале триаса по сравнению с поздней пермью резко снижается фитогеографическая дифференциация. Нечеткая выраженность географической дифференциации экваториальной, сибирской и гондванской флор среднего – позднего триаса, а так же широкое распространение некоторых родов свидетельствует о нерезкости климатических зон. Видимо, по всей Земле был безморозный климат. Для растительности триаса характерно интенсивное развитие различных групп голосеменных: хвойных, гинкговых и цикадовых, определивших мезофитный облик флоры. Изменение флоры на рубеже перми и триаса происходило постепенно и растянулось на многие миллионы лет. В разных регионах мира смена палеофитных комплексов на мезофитные (т.е. группы, которым предстоит доминировать в мезозойских флорах, точнее, от средины триаса до середины мела) отмечается на разных стратиграфических уровнях и часто не совпадает с границей перми и триаса, проводимой по фауне. Например, на территории Восточной Европы доминирование мезофитных элементов отмечается задолго до конца перми. Детальная реконструкция этого перехода невозможна из-за неполноты стратиграфических данных и наличия крупных перерывов в разрезах. В триасе остаются достаточно многочисленными такие палеофитные растения, как папоротники и хвощи. К концу триаса выделяется три основных фитогеографических зоны – Австралийская, Евро-Синайская и СибирскоКанадская.
В триасовых отложениях впервые появляются примитивные Hymenoptera (пилильщики), Phasmatodea (палочники), Diptera (двукрылые), однако высокая степень дифференциации этих групп, вероятно, указывает на их более раннее возникновение.
Для животного мира суши и пресноводных бассейнов характерно возрастающее значение рептилий и сокращение амфибий – стегоцефалов. Среди последних были достаточно распространены представители родов Benthosuchus в раннем и Mastodonsaurus – в позднем триасе. Вымирали обычные для перми рептилии - зверообразные, котилозавры; их сменили новые группы: в среднем триасе появились первые ящеротазовые динозавры (хищные и растительноядные). В конце триаса происходит важное событие, отразившееся на всем характере развития органического мира – появление первых млекопитающих (Prototheria – Первозвери). Это были небольшие, размером с крысу, яйцекладущие животные, питавшиеся насекомыми или мелкими позвоночными и вскармливающие детенышей молоком. Происхождение млечных желез у древних млекопитающих связывают с видоизменением потовых желез. В связи с этим Д. Холден высказал предположение, что первоначальной функцией млечных желез было не столько выкармливание, сколько "выпаивание" детенышей, т.е. снабжение их необходимой влагой и солями. На самых ранних этапах эволюции млекопитающих начинается и расхождение основных эволюционных стволов, ведущих, с одной стороны, к однопроходным (Monotremata), с другой к сумчатым (Marsupialia) и плацентарным (Placentalia). Вымершие и современные отряды яйцекладущих (отряд однопроходные – Monotremata; утконосы, ехидны), возможно, представ-
80