

Paleo / MARTYNOV
.pdfраспространение покрытосеменных отмечается уже в середине периода). Остается не ясным и ключевой вопрос об одновременности процессов вымирания на разных континентах и в разных океанах и морях. Например, по данным Р. Слоуна, в Северной Америке динозавры (трицератопсовая фауна) существовали еще несколько миллионов лет в начале палеогена после их вымирания в других регионах.
Рассматривая причины вымирания, некоторые ученые слишком много внимания уделяют взаимоотношениям организмов, в частности вытеснению более совершенными организмами менее совершенных. Например, аммонитов с их громоздким гидростатическим аппаратом – полой камерной раковиной – могли вытеснить, истребляя в большом количестве, более активные и подвижные рыбы. Тем не менее, имеющие сходный гидростатический аппарат представители отряда Nautilida, появившись в девоне, дожили до настоящего времени – Nautilus, хотя никогда не достигали такого пышного развития, как аммоноидеи, и всегда занимали довольно скромное место в историческом развитии биосферы. Роковую роль в судьбе динозавров могли сыграть мелкие млекопитающие, пожирая яйца этих животных. Однако млекопитающие возникли еще в позднем триасе и на протяжении примерно 130 млн. лет, прошедших до конца мезозоя, оставались малозаметной и малозначительной группой.
Безусловно, взаимоотношения организмов играли огромную роль в эволюционном процессе. Однако совпадение этапов развития многих как наземных, так и морских организмов приводит к выводу о том, что в процессе эволюции на рубеже мезозоя и кайнозоя решающее значение имели какие-то общепланетарные изменения абиотических условий. Очень часто кризис в эволюции связывают с изменением климата, тектоническими движениями, изменявшими физико-географические условия, вулканизмом. Например, французский палеонтолог Л. Гинзбург считает, что в результате поднятия материков к концу мела произошла значительная регрессия моря. В ходе этой регрессии площадь эпиконтинентальных морей уменьшилась примерно в 50 раз. Именно мелкие эпиконтинентальные моря и предоставляли наиболее благоприятную среду для обитания морских организмов. Безусловно, значительное сокращение акватории морей должно было негативно сказаться на самых различных группах морских организмов, но остается непонятным избирательность вымирания. Почему вымерли морские рептилии, моллюски рудисты, аммониты, белемниты, многие наутилоидеи и т. д., но совершенно не пострадали, например, костистые и пластинчатожаберные рыбы. В последнее время высказываются различные гипотезы о космических причинах экологической катастрофы на рубеже мела и палеогена, например, взрыв сверхновой звезды в относительной близости от Солнечной системы (порядка 100 световых лет). По мнению авторов этой гипотезы, повышение интенсивности космического излучения должно было вызвать резкое усиление атмосферной циркуляции с образованием вихрей, результатом чего может быть резкое (хотя геологически кратковременное) общее понижение температуры на поверхности Земли. Этот фактор и сыграл роковую роль в вымирании ди-
101

нозавров, приспособленных к теплому, ровному климату мезозоя. В этом случае нерешенным остается вопрос, почему не пострадали растения, также достаточно чувствительные к климатическим колебаниям.
Недавно получены новые интересные данные относительно причины великих перемен органического мира в конце мезозоя. В местах непрерывного осадконакопления пограничных слоев мела и палеогена, таких как моря, глубоководные океанские впадины и на континентах на границе маастрихтского и датского ярусов обнаружено значительное повышение содержания платиноидов – иридия, осмия и др. Полагают, что эти элементы привнесены на Землю в результате падения астероида диаметром не менее 10 км – крупного метеорита или кометы 65 млн. лет назад. Подземный кратер на месте падения этого астероида диаметром 180 км найден на п-ове Юкатан. Многие ученые считают, что вымирание животных и растений в конце мезозоя было связано с космической катастрофой, резко нарушившей физико-химические условия среды (температуру, облачность, повышенное содержание аэрозолей в атмосфере и т. д.). Однако вымирание организмов не было одновременным. Многие преобразования в органическом мире происходили длительное время и осуществлялись задолго до предполагаемой катастрофы. Некоторые группы организмов миновали рассматриваемый рубеж без особых изменений, у других групп перемены осуществлялись в иное время. И тем не менее, вымирание организмов на рубеже мела было весьма существенным.
* Обзор всех гипотез, выдвинутых в связи с мезозойским вымиранием, потребует отдельного монографического исследования. Кратко их можно сгруппировать по следующим признакам: 1) тектонические гипотезы, согласно которым причина вымирания лежит в изменении конфигурации континентов и океанов, в распределении суши и моря, обусловленных тектоническими движениями; 2) гипотезы газовых факторов – вымирание происходит вследствие изменения соотношения газов в атмосфере; 3) гипотезы климатических изменений – изменения климата являются причиной глобального кризиса; 4) эвстатические гипотезы – причина лежит в колебании уровня океанов, грандиозные трансгрессии и регрессии (наступления и отступления) моря; 5) гипотезы изменения солености, которая и является основой массового вымирания; 6) вымирания - результат резких изменений количества микроэлементов; 7) вымирание является следствием воздействия радиоактивных элементов; 8) повышение температуры, как следствие падения огромных метеоритов; 9) причина массовых вымираний – ультрафиолетовая радиация; 10) космические факторы – например, взрыв сверхновой звезды; 11) динозавры вымерли от отравления алкалоидами содержащимися в покрытосеменных растениях; ее не менее оригинальный вариант - от запоров; 12) вымирание организмов является следствием израсходования "эволюционной энергии"; 13) "старение видов"; 14) конкуренция с млекопитающими; 15) эпидемии; 16) смещение межпозвонковых дисков, вызванное громадным весом и т. д. и т. п.
102
ИСТОРИЯ ЗЕМЛИ В КАЙНОЗОЙСКУЮ ЭРУ (ЭРАТЕМА, ГРУППА)
(греч. kainos - новый; zoe - жизнь)
ОБЩИЕ СВЕДЕНИЯ
Кайнозойская, или новая эра геологической истории Земли включает и современную эпоху развития. Начало 65 млн. лет назад (начиная с датского века), полная продолжительность неясна, по некоторым данным может достигнуть 90 млн. лет. Кайнозойскую эру подразделяют на три периода: палеогеновый, неогеновый и четвертичный (антропогеновый). В ряде стран продолжают применять старое деление на два периода (третичный и четвертичный), которое было принято в 1881 г на 2-й сессии Международного геологического конгресса в Болонье, где одновременно было утверждено деление третичной системы на два отдела - палеогеновый и неогеновый. Еще в конце XIX в. некоторые геологи высказали мнение о том, что палеоген и неоген следует считать самостоятельными периодами, учитывая значительные различия в составе органического мира, физико-географической обстановке и геологической истории. Хотя по этому поводу решение Геологического конгресса не принято, во многих странах кайнозойскую эру делят на три периода. Общепринятые подразделения для палеогена, неогена и антропогена не выработаны.
ОРГАНИЧЕСКИЙ МИР
Кайнозойская эра знаменует новый этап в развитии органического мира. Характерной особенностью органического мира кайнозоя является необычайный расцвет покрытосеменных растений, млекопитающих и птиц на суше, двустворок и гастропод в море. Многие из характерных для мезозоя морских и наземных животных вымерли или утратили свое значение к началу палеогена (изменение флоры произошло раньше, в течение раннего мела). Среди морских беспозвоночных исчезли аммоноидеи, белемниты, иноцерамы, а в других группах изменился родовой и видовой состав. Среди позвоночных вымерли наземные динозавры, летающие птерозавры, гигантские морские рептилии; их место заняли быстро эволюционировавшие млекопитающие.
В морях с начала палеогена стали возникать и быстро распространяться новые роды и семейства двустворок, гастропод и костистых рыб. На суше доминировали млекопитающие, птицы и насекомые, продолжалось развитие покрытосеменных растений. Углеобразование наблюдалось преимущественно в неогене.
ПАЛЕОГЕНОВЫЙ ПЕРИОД (СИСТЕМА)
Палеогеновый период начался 65 млн. лет назад и закончился 25 млн. лет назад, его продолжительность 40 млн. лет. Как самостоятельное подразделе-
103
ние был впервые выделен К. Науманном в 1866 г. В геохронологии палеогена главную роль играют биостратиграфические зоны, устанавливаемые по микроорганизмам (планктонным фораминиферам, нанопланктону) и нуммулитам (Табл. 13).
В раннем палеогене произошло расхождение Австралии и Антарктиды. Индийский массив соединился с Евразией, вызвав образование горных систем Памира и Гималаев. Сближение Африканского блока с Европой сузило акваторию океана Тетис до размеров Средиземного моря. В течение этого периода осуществился ряд фаз альпийского горообразования, сформировались осевые хребты современных Атласа, Пиренеев, Альп, Карпат, Крыма, Кавказа, Копетдага, Памира и Гималаев. Климат в раннем и среднем палеогене оставался теплым и влажным, только к концу этого периода произошло общее понижение температуры и заметная аридизация климата.
Таблица 13
Геохронологическое (стратиграфическое) подразделение палеогеновой системы
Эпоха (отдел) |
Подотдел |
Век (ярус) |
Олигоцен |
Верхний |
Хаттский P3 h |
(верхний) |
Нижний |
Рупельский P3 r |
|
|
Латторфский P3 l |
Эоцен |
Верхний |
Приабонский P2 p |
(средний) |
|
Бартонский P2 b |
|
Средний |
Лютетский P2 l |
|
Нижний |
Ипрский P2 I |
Палеоцен |
Верхний |
Танетский P1 t |
(нижний) |
|
Монтский P1 m |
|
Нижний |
Датский P1 d |
ОРГАНИЧЕСКИЙ МИР
Органический мир моря палеогенового периода был очень разнообразен. Среди беспозвоночных господствовали двустворчатые и брюхоногие моллюски, фораминиферы, широкое распространение получили морские ежи, кораллы, губки, мшанки, остракоды, а из растений – золотистые и диатомовые водоросли. Наиболее важными для стратиграфии и хорошо изученными являются моллюски, фораминиферы, морские ежи, остракоды и золотистые водоросли – кокколитофориды.
Простейшие – фораминиферы получили исключительное развитие в палеогене, в большом количестве были представлены как планктонные, так и бентосные формы. Важнейшими среди планктонных фораминифер были ро-
ды Globigerina, Subbotina, Globorotalia, Acarinina, Hantkenina, а среди бен-
тосных крупных фораминифер (с диаметром раковины до 15 см) расцвета
104
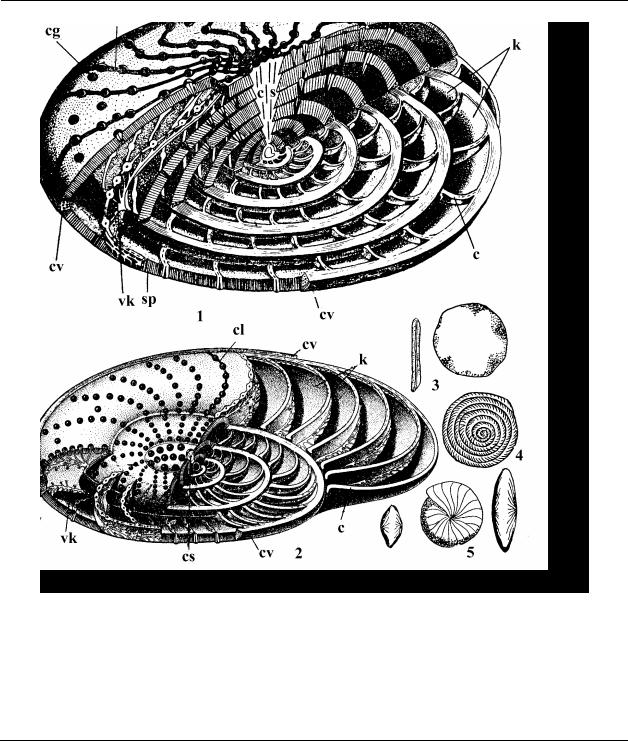
достигают представители отряда Nummulitida - роды Nummulites, Assilina, Operculina, Discocyclina, которые быстро эволюционировали, получили широкое развитие в эоцене, к началу олигоцена большинство из них вымерло (Рис. 44). Эти формы обитали на небольшой глубине в теплых морях, входивших в состав Тетиса. Нуммулиты были породообразующими организмами, и сформировали мощные толщи нуммулитовых известняков (например, в Египте их мощность достигает 400 м), в связи с чем палеоген иногда называют нуммулитовой системой. Достаточно широко были распространены радиолярии.
Рис. 44. Тип Sarcomastigophora, Класс Foraminifera – Фораминиферы, Отряд Nummulitida: 1, 3, 4, 5 – Nummulites (палеоген): 1 – схема строения раковины, продольное сечение; 3, 5 – внешний вид; 4 – экваториальное сечение; 2 – схема строения раковины рода Operculina; vk – внутрисептальные каналы, k – камеры, с – септы, cv – спиральный валик, cg – септальные гранулы, cl – септальная линия, sp – стенка раковины, cs – центральный столбик
105
Двустворчатые и брюхоногие моллюски достигли в палеогене исключительного разнообразия. В отличие от других классов моллюсков, они обитали не только в морях с нормальной соленостью, но и в солоноватоводных и пресноводных бассейнах, а брюхоногие к тому же были широко распространены на суше. Число их родов и видов чрезвычайно велико. Многие из палеогеновых родов моллюсков возникли еще в конце мелового периода, но особенно много их появилось в эоцене. На границе эоцена и олигоцена из-за похолодания значительно изменились родовые комплексы (олигоценовый комплекс моллюсков очень близок к неогеновому и современному). Среди двустворок следует отметить: Area, Chlamys, Crassatella, Ostrea, Glycymeris, Spondylus; среди гастропод – Turritella, Cerithium, Pleurotoma и Natica (Рис. 45).
Теплые воды палеогеновых морей нормальной солености были благоприятны для широкого распространения морских ежей: Echinocorys, Clypeaster, Conoclypeus и др, шестилучевых кораллов, которые вместе с другими организмами участвовали в формировании коралловых рифов. Часто встречались губки. Из членистоногих были обычны остракоды, которые обитали как в морских, так и в пресноводных водоемах (наиболее характерны
Trachyleberis, Cytheridea, Loxoconcha). Менее распространены десятиногие ракообразные, в частности, пресноводные и морские раки, креветки. Мшанки и брахиоподы играли незначительную роль.
Применение электронной микроскопии позволило установить широкое распространение в породах палеогена кокколитофорид. В последние годы установлено, что они играли весьма важную роль в палеогене, лишь кажущуюся меньшей, чем в позднем мелу. Расцвет кокколитофорид был в эоцене, в эту эпоху они являлись породообразующими организмами. Из морских породообразующих микроскопических водорослей в палеогене также получили пышное развитие диатомовые и кремневые жгутиковые. Состав диатомовых определялся соленостью, температурой и гидродинамическим режимом бассейнов, в связи с чем они активно используются для реконструкции физикогеографических условий прошлого.
Среди морских позвоночных, как и в позднем мелу, господствовали костистые рыбы. Морская ихтиофауна палеогена включала все ныне существующие крупные группы рыб, хотя видовой состав отличался от современного. В палеогеновых морях появились водные млекопитающие, которые заняли освободившиеся экологические ниши морских рептилий. Это были древнейшие китообразные (Cetacea), остатки которых отмечены со среднего эоцена. По всей видимости, китообразные возникли от каких-то примитивных наземных хищников – креодонтов (Carnivora, Creodonta), перешедших к рыбоядности и к амфибиотическому, а затем и чисто водному образу жизни. Пропорции тела архаических китов (Archaeoceti) заметно отличались от современных групп, их туловище было длинное и относительно тонкое (у наиболее крупной формы зейглодона – Zeuglodon - достигало в длину 21 м при диаметре не более 2,5 м).
106

Рис. 45. Характерные ископаемые останки палеогеновых беспозвоночных: класс Foraminifera – Фораминиферы, 1-3 – Nummulites (палеоген – ныне, преимущественно эоцен); 4 – Assilina (палеоцен – эоцен); 5 – Operculina (поздний мел – ныне). Класс Bivalvia – Двустворчатые моллюски, 6 – Cardium (неоген – ныне); 7 – Spondylus (юра – ныне, преимущественно кайнозой); 8 – Chlamus (триас – ныне); 9 – Glycymeris (мел – ныне, преимущественно кайнозой). Класс Gastropoda – Брюхоногие моллюски, 10 – Turritella (мел – ныне); 11 – Cerithium (поздний мел – ныне); 12 – Pleurotoma (палеоген – ныне); 13 – Nassarius
(эоцен – ныне); 14 – Natica (палеоген – ныне). Класс Echinoidea – Морские ежи, 15 –
Conoclypeus (эоцен – миоцен); 16 – Clypeaster (эоцен – ныне)
107
Примитивные представители современных групп китообразных – зубатых (Odontoceti) и беззубых (Mystacoceti) китов – появились, соответственно, в позднем эоцене и олигоцене.
Органический мир суши. Наземная фауна палеогена сильно отличалась от мезозойской. Пресмыкающиеся были представлены немногочисленными крокодилами, ящерицами, черепахами и змеями; земноводные – гигантские саламандры, лягушки и жабы – не играли заметной роли среди позвоночных палеогена. Подлинного расцвета достигают птицы, став самой многочисленной по числу видов группой наземных позвоночных. Между птицами и млекопитающими произошел своеобразный "раздел сфер влияния". Большинство видов млекопитающих, возникших от сумеречных и ночных мезозойских форм, сохранило и в кайнозое предпочтение к ночной активности, тогда как птицы, потомки гелиотермных рептилий, остались, до настоящего времени, преимущественно дневными животными. После вымирания динозавров птицы, как и млекопитающие, предпринимают попытку занять освободившиеся экологические ниши крупных хищников. Так, в раннем эоцене появляются крупные наземные нелетающие птицы с редуцированными крыльями. Например, североамериканская эоценовая диатрима (Diatryma – "ужасный журавль") достигала высоты 2 м, обладала массивным клювам с острыми краями и габитусом напоминала хищных динозавров. Неудача птиц в этой роли может быть связана с несовершенством (по сравнению с млекопитающими) размножения. Во всяком случае "пернатые динозавры" вымерли после появления высших групп хищных млекопитающих.
Вымирание динозавров оставило свободным широкий спектр адаптивных зон, которые стали быстро заполняться териевыми млекопитающими (Theria
– Звери). Фауна млекопитающих в начале периода (палеоцен) оставалась по существу той же, что и в позднемеловое время. В ней были представлены только группы, возникшие еще в мезозое. Это были мелкие формы, напоминавшие многие ранние группы насекомоядных и всеядных сумчатых Южной Америки. Активно развиваясь, они начинают господствовать на суше и достигают большого разнообразия уже к среднему палеоцену, известно по крайней мере 30 отдельных семейств. К сожалению, палеонтологическая летопись позволяет проследить родственные связи только для очень немногих групп. В эоцене уже существовали все основные отряды кайнозойских мле-
копитающих: грызунов (Rodentia), хищных (Carnivora, Creodonta), непарно- (Perissodactyla) и парнокопытных (Artiodactyla), появились первые немногочисленные насекомоядные (Insectivora) и зайцеобразные (Lagomorpha). Быстрая адаптивная радиация новых групп растительноядных привела к вымиранию архаичных форм – многобугорчатых (средина эоцена), кондиляртр (Condylarthra – конец эоцена) и диноцерат (Dinocerata – страшнорогие). Своеобразным убежищем для некоторых примитивных групп млекопитающих стала Южная Америка, которая к концу раннего эоцена обособилась от Северной Америки и оставалась изолированной до плиоцена. В палеогене и неогене здесь существовал удивительный мир причудливых форм, сохранивших общий примитивный уровень организации (подробнее фауна
108

Ю. Америки и Австралии будет рассмотрена ниже).
Среди высших копытных раньше других начали адаптивную радиацию непарнокопытные, которые уже в эоцене были представлены большим разнообразием форм. Центром эволюции этого отряда была Северная Америка (в течение всего кайнозоя соединялась с Азией через область Берингова пролива)*.
Рис. 46. отряд Непарнопалые –
Perissodactyla. Индрикотерий
(Indricotherium) реконструк-
ция внешнего вида (ср. олигоцен)
Непарнокопытные пережили максимальный расцвет в палеогене. В олигоцене достигли огромных размеров древние носороги, среди которых встречаются гиганты – безрогие индрикотерии
(Indricotheriidae), они были распространены в Азии и обитали в лесостепях и саваннах. Это были самые крупные млекопитающие, обитавшие на суше (по длине и массе тела они уступали только динозаврам): длина их тела достигала 7 – 8 м, высота в холке – до 5,5 м. Довольно длинная шея могла поднять огромную голову (череп имел длину до 1,2 м) на высоту около 8 м над землей (Рис. 46). Постепенная аридизация климата в азиатских степях привела к вымиранию индрикотериевой фауны в начале неогена.
Адаптивная радиация парнокопытных началась несколько позже, чем непарнокопытных, однако уже в олигоцене существовали примитивные представители современных семейств свиных (Suidae), верблюдов (Camelidae), оленьков (Tragulidae) и целый ряд вымерших групп, их эволюция происходила, в основном, в неогене.
Приматы – типичные млекопитающие кайнозоя – появились, вероятно, в конце мезозоя. В начале палеоцена уже существовали лемуры – наиболее примитивная группа среди современных приматов, называемых полуобезьянами. В конце эоцена появились настоящие обезьяны – антропоиды (Anthropoidea), древнейшие остатки которых известны из нижнего олигоцена Египта. В начале олигоцена наиболее примитивными были парапитеки (Parapithecus) - возможные предки более высокоразвитых антропоидов.
В Африке развивалась своя оригинальная фауна растительноядных млекопитающих. В состав этой фауны входили крупные хоботные (Proboscidea), их расцвет наступил в неогене, даманы или жиряки (Embrithopoda), и перешедшие к водному образу жизни сиреновые (Sirenia), у которых развился
* Несмотря на наличие эндемичных групп, фауна млекопитающих Азии, Европы и Северной Америки была в целом сходной на протяжении всего кайнозоя, что является следствием практически непрерывного обмена фаунами в течение кайнозоя.
109
хвостовой плавник, передние конечности преобразовались в ласты, а задние редуцировались.
Древнейшие представители плацентарных хищников (Carnivora) – креодонты (Creodonta) известны с раннего палеоцена, и возникли, вероятно, в Северной Америке – по некоторым чертам организации были близки к примитивным копытным кондиляртрам и, вероятно, имели общее с ними происхождение. Обособление эволюционного ствола хищников произошло, вероятно, еще в конце мелового периода. Наиболее ранние формы креодонтов имели небольшие размеры, тонкие, короткие конечности и стройное тело, представляя переходной тип от насекомоядных предков. Уже в палеоцене появляются крупные хищники, например, арктоцион (Arctocyon), размерами и телосложением напоминавших современных медведей. В раннем олигоцене существовали еще более крупные формы, например, эндрюсархус (Andrewsarchus) достигал в длину 4 м при высоте в холке до 2 м. Поведение креодонтов, вероятно, оставалось достаточно простым в сравнении с современными хищниками как в приемах охоты, так и в уходе за потомством. Уже из среднего палеоцена известны остатки представителей высших хищников (Fissipedia). К концу эоцена от примитивных родов могли начать дивергировать современные семейства. Большинство послеэоценовых хищных можно отнести к ныне живущим семействам. В олигоцене последние креодонты вымирают.
Среди наземной флоры продолжалось развитие покрытосеменных растений, чему способствовал теплый климат палеогена. В Европе, до берегов Балтийского моря, произрастали густые тропические и субтропические леса с вечнозелеными пальмами, магнолиями, миртами, фикусами; из хвойных - гигантские секвойи, араукарии, кипарисы; в лесной тени – папоротники; по морскому побережью – мангровая растительность. Эта флора в Евразии называется полтавской. Подобная флора существовала в Америке, Африке и в Азии. Листопадные деревья – дуб, бук, каштан, береза, тополь и некоторые хвойные преобладали на севере Европы (включая Гренландию и Шпицберген), на большей части Азии и на севере Америки – тургайская флора. На всей этой территории произрастали также отдельные представители субтропической растительности. Климат здесь был близок к современному климату Средиземноморья, Калифорнии.
В палеогене отчетливо выделяются три этапа обновления покрытосеменных: палеоценовый, эоценовый и олигоценовый, что связано с постепенным похолоданием и общей эволюцией растений. Теплолюбивая флора Евразии и Северной Америки постепенно смещалась к югу, сменяясь в умеренном поясе листопадными и хвойными лесами. Кроме похолодания на обширных площадях происходила аридизация климата. На огромной территории Центральной Азии это привело к постепенному исчезновению лесов и развитию лесостепей, степей и саванн. В олигоцене здесь преобладали злаки, а деревья и кустарники сохранились в долинах рек.
Состав и расселение органического мира в палеогене свидетельствуют о существовании зоо- и фитогеографических провинций. По морским беспо-
110