

Paleo / MARTYNOV
.pdfИСТОРИЯ ЗЕМЛИ В ПАЛЕОЗОЙСКУЮ ЭРУ (ЭРАТЕМА, ГРУППА)
(греч. palaios – древний; zoe - жизнь)
ОБЩИЕ СВЕДЕНИЯ
Палеозой – эра древней жизни, начинает новый крупный эон в истории Земли – фанерозой. Впервые была выделена в 1837 г английским геологом Седжвиком. Палеозойская эра – наиболее продолжительная эра фанерозоя, она длилась 335 млн. лет и подразделяется на шесть периодов: кембрийский, ордовикский, силурийский, девонский, каменноугольный и пермский. В течение палеозойской эры земная кора претерпела значительные преобразования, выразившиеся в изменении конфигурации морских бассейнов и суши, в появлении и исчезновении наземных и подводных горных хребтов, в изменении положения полюсов и соответственно климатических поясов. Многие геологи считают, что до палеозоя существовал единый континентальный блок – Пангея – и единый протоокеан. В дальнейшем единый протоконтинент распадается на два суперконтинента: северный – Лавразию и Южный – Гондвану (современные Ю. Америка, Африка, о. Мадагаскар, юг Азии, Австралия, Антарктида), между которыми простиралась морская внутренняя впадина Тетис. На все эти физико-географические преобразования чутко реагировал органический мир Земли. Именно изменения в мире древних животных и растений послужили основой для выделения палеозойской эры, и ее отдельных периодов. По этим же признакам палеозойская эра может быть разделена на две части (подэры) - ранний (кембрий, ордовик, силур) и поздний (девон, карбон, пермь) палеозой. Двучленное деление палеозоя подтверждается в первую очередь историей развития крупных таксономических единиц ранга классов. Большинство новых классов появилось в кембрии и девоне, а к силуру и перми число их уменьшилось.
Иногда палеозой делят на три части: нижний (кембрий - ордовик), средний (силур - нижний карбон), верхний (средний карбон – пермь). Мы будем придерживаться двучленного деления палеозоя.
ИСТОРИЯ ЗЕМЛИ В РАННЕМ ПАЛЕОЗОЕ ОРГАНИЧЕСКИЙ МИР
Органический мир раннего палеозоя характеризуется становлением и развитием всех типов животных и низших растений. Его своеобразие состоит в том, что организмы обитали, главным образом, в морях, а суша была слабо освоена растениями, и тем более животными. В морях господствовали беспозвоночные, представленные древними группами, и разнообразные водоросли. Обитавшие в морских водоемах позвоночные были примитивны и немногочисленны. На суше, вероятно, в течение долгого времени с начала кембрия, существовали только примитивные водоросли и бактерии. Лишь во второй половине этого этапа на суше появились высшие растения и назем-
31
ные животные из типа членистоногих. С деятельностью растений связано накопление в атмосфере кислорода, а значит, и озона, который создает экран, не пропускающий к поверхности Земли ультрафиолетовое излучение Солнца. К концу раннего палеозоя количество кислорода в атмосфере достигло 2% (так называемая точка Беркнера-Маршалла), т. е. уровня, необходимого для создания защитного слоя.
КЕМБРИЙСКИЙ ПЕРИОД (СИСТЕМА)
По данным ядерной геохронологии кембрийский период начался 570 млн. лет назад и закончился 490 млн. лет назад, его общая продолжительность – около 80 млн. лет (Табл. 3).
Кембрийская система была выделена в 1835 г на территории Великобритании А. Седжвиком и получила название от древнего наименования Уэльса
– Cambria. В начале периода суша была представлена несколькими материками, расположенными в основном в Южном полушарии: Гондваной (включающей Южно-Американскую, Африканскую, Антарктическую, Австралийскую платформы, а также Индостан и Китай), Лаврентией (СевероАмериканская платформа), Балтикой (Европейский щит), Сибирью и Казахстаном. Их омывал единый океан Панталасса. Лаврентия и Балтика были разделены океаном Япетус. Климат кембрия в целом был жаркий.
Таблица 3
Геохронологическое (стратиграфическое) подразделение кембрийской системы
Эпоха |
Надъярус |
Век (ярус) |
(отдел) |
|
|
Верхний |
|
Батырбайский Є3 b |
|
|
Аксайский Є3 ak |
|
|
Сакский Є3 s |
|
|
Аюсаканский Є3 as |
Средний |
|
Майский Є2 m |
|
|
Амгинский Є2 am |
Нижний |
Ленский Є11 |
Тойонский Є1 tn |
|
|
Ботомский Є1 b |
|
Алданский Є1а |
Атдабанский Є1 at |
|
|
Томмотский Є1 t |
|
ОРГАНИЧЕСКИЙ МИР |
|
Кембрий начинает эру палеозойской жизни. Уже в отложениях нижнего кембрия найдены остатки почти всех типов животных, известных и в настоящее время, в том числе и хордовых и, что особенно интересно и важно, многие из них приобрели способность строить прочный, сначала хитиново-
32
фосфатный, а затем и известковый наружный или внутренний скелет. Таким образом, к кембрию приурочен важный этап в развитии жизни на Земле – появление у животных твердого скелета.
Основу фауны периода составляли археоциаты, трилобиты и многие представители кишечнополостных, червей, моллюсков и некоторых других групп животных. Все это – водные и почти исключительно морские формы.
Рис. 5. Тип Chordata –
Хордовые, п/т Acrania – Бесчерепные. Pikaia – первое известное хордовое, сланцы Берджес (средний кембрий)
Кембрий – время бурного расцвета членистоногих, и особенно
одной из групп – трилобитов. Они были наиболее массовыми животными и составляли около 60% всей фауны периода. Трилобиты – морские членистоногие, выделяемые в отдельный, полностью вымерший подтип Trilobitomorpha (известно более 10 000 видов), в составе которого описано около 10 классов, представленных только в раннем кембрии почти 50 семействами. Наиболее многочислен и разнообразен в палеозое был класс Трилобиты (Trilobita), в пределах которого по числу сегментов туловищного отдела выделяют два подкласса – малочленистые и многочленистые. Для этого периода характерны малочленистые (подкласс Miomera) трилобиты (Agnostus, Pagetia и др.), которые почти полностью исчезли к концу кембрия (Рис. 7: 3- 4). Многочленистые (подкласс Polymera) достигли значительного разнообразия, но почти все характеризуются слабым (по сравнению с цефалоном) развитием пигидия, большим числом туловищных сегментов, опистопарными лицевыми швами, хитиново-фосфатным панцирем и отсутствием способно-
сти свертываться в шарик (Redlichia, Olenellus, Paradoxides и др.) (Рис. 7: 7- 9). Трилобиты вели донный образ жизни, зарывались в толщу осадков, ползали по дну, но имелись и активно плавающие пелагические формы. Это были первые из известных нам животных с высокоразвитым зрением. Глаза трилобитов имели от нескольких десятков до 15 000 фасеток, хотя известны и слепые формы. Среди них, видимо, были как растительноядные формы, так и илоеды, падальщики, активные хищники. Предполагают, что некоторые трилобиты, возможно, охотились даже на своих сородичей. Крупнейшие из трилобитов имели длину свыше 70 см, а самые маленькие не достигали и сантиметра. Индивидуальное развитие трилобитов является ярким подтверждением правильности выдвинутой А.А. Захваткиным теории происхождения метаморфоза у насекомых. Он считал, что исходным для Arthropoda был первично-личиночный тип развития (анаморфоз), характеризующийся плавным преобразованием организации. Так, у трилобитов из яйца вылуплялась
33
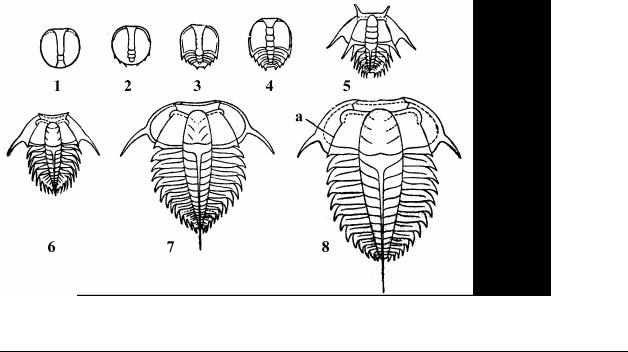
личинка-протаспис с четырьмя ларвальными сегментами (Рис. 6: 1). В ходе дальнейшего развития, после каждой линьки появлялись новые, постларвальные туловищные сегменты, зона роста находилась перед пигидием (Рис. 6: 3-7). Анаморфоз характерен и для современных веслоногих раков – циклопов.
Рис. 6. Стадии индивидуального развития трилобитов от личинки (1) до стадии взрослого организма (8): 1 – 4 – ранние стадии развития; 5 – 7 – поздние стадии развития; 8 – взрослый трилобит Leptoplastus salteri, а – лицевой шов
Трилобиты в кембрии быстро эволюционировали, в связи с чем стратиграфия кембрийской системы основана почти исключительно на их остатках.
Наряду с трилобитами в кембрии в различной степени были развиты представители всех остальных подтипов членистоногих, среди которых особенно следует отметить ракообразных (остракод и филлокарид).
Другой важной группой, характерной исключительно для кембрийской фауны, были археоциаты. Полностью вымерший тип Археоциаты (Archaeocyathi) – появившись в начале кембрия, к средине раннего кембрия достигает расцвета, но до среднего кембрия доживают только отдельные представители. Археоциаты населяли теплые мелководные моря и составляли около 30% фауны периода.
Внешним видом, строением и биологией они сильно похожи на современных губок, их тело имело вид кубка различной формы (Рис. 7: 10, 11). По способу питания они были фильтраторами и составляли основную часть неподвижного бентоса раннего кембрия. Обычно они прикреплялись к дну (при помощи каблучка), хотя могли быть и свободнолежащими. Диаметр кубков – от нескольких миллиметров до 70 см, высота от нескольких сантиметров до 1,5 метра. Форма кубка была чрезвычайно разнообразна, при этом изменчивость носила и внутривидовой характер, поскольку сильно зависела от гидрологического режима водоема – скорости течения, температуры, что свидетельствует о низком уровне организации. В раннем кембрии археоциаты достигли большого разнообразия.
34
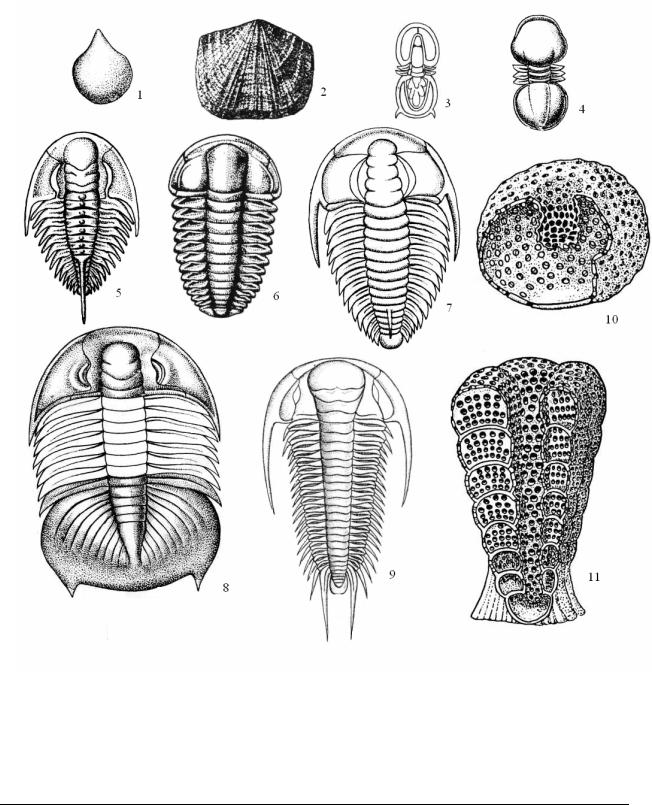
Рис. 7. Характерные ископаемые кембрийского периода: Тип Brachiopoda - Брахио-
поды: 1 – Mickwitzia monilifera; 2 – Billingsella fluctuosa; п/т Trilobitomorpha - Трилобиты, класс Miomera: 3 – Agnostus pisiformis; 4 – Pagetiellus lenaicus. Класс Polymera: 5 – Shmidtiellus mickwitzi; 6 – Ellipsocephalus hoffi; 7 – Redlichia chinensis; 8 – Dikelocephalus minnesotensis; 9 – Paradoxides bohemicus. Археоциаты – Archaeocyathi: 10 – Capsulocyathus; 11 – Coscinocyathus
Они были представлены простыми одностенными и сложными двустенными формами как одиночными, так и колониальными. Хотя мы не имеем сведений о строении мягкого тела археоциат, они совместно с губками включаются в надраздел Parazoa. По строению интерваллюма и характеру онтогенеза тип разделяется на два класса: правильные (Regulares) и неправильные (Irregulares). Археоциаты были первыми скелетными организмами,
35

участвовавшими в построении рифов. Позднее, уже в ордовике, им на смену придут кораллы. К концу кембрия археоциаты вымерли.* Широкое географическое распространение и быстрая эволюция археоциат в течение раннего кембрия позволяет широко использовать их для расчленения и корреляции нижнекембрийских отложений.
Сравнительно широко были распространены в кембрии беззамковые брахиоподы (Тип Brachiopoda, класс Inarticulata) с хитиново-фосфатными и известковыми раковинами (Obolus, Kutorgina); в это время начинали свое развитие и замковые брахиоподы (класс Articulata). Первые представители брахиопод обнаружены в самом основании нижнего кембрия; активно развиваясь, эта группа уже в среднем кембрии становится второй по численности группой среди скелетных ископаемых. Кембрийские моря были заселены различными гидроидными и сцифоидными кишечнополостными, вместе с которыми уже в начале кембрия появились коралловые полипы. Активно развивались моллюски; если в самом начале периода этот тип животных был представлен моноплакофорами и гастроподами, то в среднем кембрии к ним присоединились двустворки, а в позднем – головоногие и полиплакофоры. Головоногие были первыми известными нам по ископаемым остаткам хищниками в морях палеозоя. На дне морей обитали многочисленные хиолиты (класс Hyolitha) – организмы, условно относимые к моллюскам.** Хиолиты встречаются с кембрия до перми, от них сохранились только конические, известковые раковины, устье которых закрывалось крышечкой (Рис. 8).
Кроме того, в кембрийских отложениях встречают остатки одноклеточных животных (однокамерных агглютинирующих фораминифер (Lukatiella) и радиолярий с простым сферическим скелетом из двуокиси кремния), губок, червей, немногочисленных, но разнообразных в систематическом отношении иглокожих (хомалозои, цистоидеи и другие древнейшие кринозои, древнейшие эхинозои), первых граптолитов – стереостолонат, с обрастающими или кустистыми полиморфными колониями.
Из отложений кембрия известны и первые остатки хордовых. Так, наиболее ранние отпечатки, возможно принадлежащие оболочникам (Tunicata), описаны из докембрийских отложений Восточной Европы (Yarnemia). К несомненным оболочникам относят фоссилии из раннего кембрия Китая (Cheungkongella), позднего кембрия Северной Америки (Palaeobotryllus).
*До недавнего времени тип считался полностью вымершим, и только в 1977 г французский исследователь Ж. Васле на экваториальных рифах Индо-Тихоокеанского бассейна находит небольшое, в несколько сантиметров диаметром, животное, которое сначала отнесли к известковым губкам. Этот организм Vaceletia crypta – имеет клеточное строение характерное для губок, в частности археоциты и хоаноциты, характеризуется лейкоидным типом ирригационной системы. Но в отличие от известковых губок имеет цельный известковый скелет, состоящих из двух стенок и горизонтальных днищ, характерный для археоциатов. По мнению ряда авторов, археоциаты являются только классом в составе типа Spongia – Губки.
** Систематическое положение класса Hyolitha до настоящего времени спорно, иногда их относят к кольчатым червям, крылоногим гастроподам, головоногим моллюскам, или выделяют в самостоятельный тип.
36

Аппендикулярии (Appendicularia) известны с раннего кембрия. Древнейшие представители бесчерепных (Acraniata) описаны из раннего кембрия Китая (Cathaymyrus) и среднего кембрия Северной Америки (Pikaia) (Рис. 5).
И, наконец, в позднем кембрии в морских водоемах уже обитали бесчелюстные рыбообразные позвоночные (Vertebrata, Agnatha). Происхождение позвоночных до настоящего времени дискуссионно. Наиболее вероятно, что они возникли в позднем протерозое – раннем кембрии от одной из ветвей древних хордовых, близких к головохордовым.
Рис. 8. Неопределенный тип, Класс Hyolitha – Хиолиты. Hyolithes acutus, вид сбоку. с – спинная сторона, b – брюшная сторона.
Гораздо богаче в кембрии становится мир низших растений, представленный синезелеными и зелеными водорослями. Достоверных остатков вегетативных органов наземных растений не известно, но как в континентальных, так и в мелководных морских отложениях кембрия обнаружены споры растений. Это дает основание некоторым авторам полагать, что наземные или полуназемные растения имелись в болотистых местах уже с самого начала кембрийского периода.
На суше в кембрийское время обитали только бактерии, грибы и, возможно, лишайники. Эти организмы являлись первыми почвообразователями. Благодаря их деятельности в конце кембрия были созданы условия для появления наземных животных. Ими стали черви (олигохеты) и члени-
стоногие (многоножки).
В завершении раздела хотелось бы остановиться на вопросе “загадочных организмов” или палеонтологических проблематик. Палеонтологу довольно часто приходится иметь дело с объектами, систематическая принадлежность которых к той или иной группе организмов неясна и более того – даже не всегда доказуема вообще их принадлежность к остаткам организмов. Все подобные остатки принято называть проблематикой (от гр. problematicos - затруднительный, спорный).
Наиболее известной и крупной группой является тип археоциат. В настоящее время достаточно хорошо разработана систематика этого типа, установлены классы, отряды и значительное число систематических единиц более низкого ранга. Выявлены гомологические ряды развития отдельных таксонов. И тем не менее, археоциаты остаются загадкой. До настоящего времени не решен вопрос о принадлежности их к многоклеточным или одноклеточным, к животным или растениям.
Совершенно загадочны и своеобразные кембрийские организмы, найденные в 1909 г американским палеонтологом Ч.Д. Уолкоттом (D. Wallcott) в глинистых сланцах (Burgess Shale) Канадских Скалистых гор. Предпола-
37
гают, что бугресские сланцы сформировались в нижнем кембрии. В них обнаружены самые разнообразные животные, отсутствующие как в более ранних, так и в последующих отложениях (Рис. 9). Среди них можно выделить чрезвычайно своеобразных членистоногих, которых нельзя отнести ни к одному из известных классов. Например, Marrella splendens – по всей видимости, бентосное животное длиной до 2 см. Сегментацией тела напоминала трилобитов, в отличие от которых имела на головной лопасти две пары длинных, направленных назад острых отростков, туловищные сегменты не имели характерных для трилобитов боковых выростов. Данный вид предлагают выделить в отдельный класс – Marrellomorpha, близкий к Trilobitomorpha. Крупная, до 20 см Sidneyia – возможно, была активно плавающим хищником (Рис. 9: 4). К членистоногим относились Habelia и Sarotrocercus (Рис
9: 2, 3).
Но если в отношении вышеперечисленных животных более или менее понятна принадлежность к конкретному типу, то целый ряд видов не имеет аналогов как среди вымерших, так и среди современных животных (Рис. 9: 5
– 11). Например, Hallucigenia – небольшое, 3 – 5 см, бентосное животное с мягким нечленистым телом и сферической головой. Передвигалась на 14 длинных нечленистых конечностях-ходулях, на спинной стороне несла 14 мягких трубок, направленных вверх. Хвостовая часть имела вид вытянутой трубки с округлым “анальным” отверстием.
Некоторые животные, похоже, обладали чертами, свойственными сразу нескольким современным типам. Так, Odontogriphus похож на сегментированного плоского червя, но по бокам от ротового отверстия были расположены сегментированные усики, как у членистоногих, а сам рот был окружен рядом конических зубоподобных выростов (Рис. 9: 9). У Nectocaris, голова и верхняя часть туловища были как у ракообразных, а нижняя часть туловища и хвост – как у позвоночных (Рис. 9: 8).
Dinomischus – были сидячими животными длинной 4 – 5 см. Их тело состояло из чашечки и стебля. Диск чашечки был окружен лепесткоподобными образованиями, плотно соединенными между собой боковыми краями. Посредине диска размещался рот, сбоку от него анальное отверстие.
В конце кембрия все вышеперечисленные кембрийские проблематики бесследно исчезают.
Совершенно очевидно, что в истории органического мира не могло быть форм “вне системы”, не связанных родством с теми или иными достаточно хорошо известными группами. Любая группа, как бы мала она ни была, должна найти свое место в системе. Выяснение этого – одна из важнейших задач исследователя, без решения которой невозможно понять эволюцию органического мира в целом и выявить закономерности этого процесса. При решении вопроса о систематическом положении таких групп теоретически возможны два решения: 1 – группы являются частью других, хорошо известных таксонов, хотя и сильно уклоняющимися их ветвями; 2 – группы принадлежат к самостоятельным высшим таксонам, как бы малы они не были по объему.
38
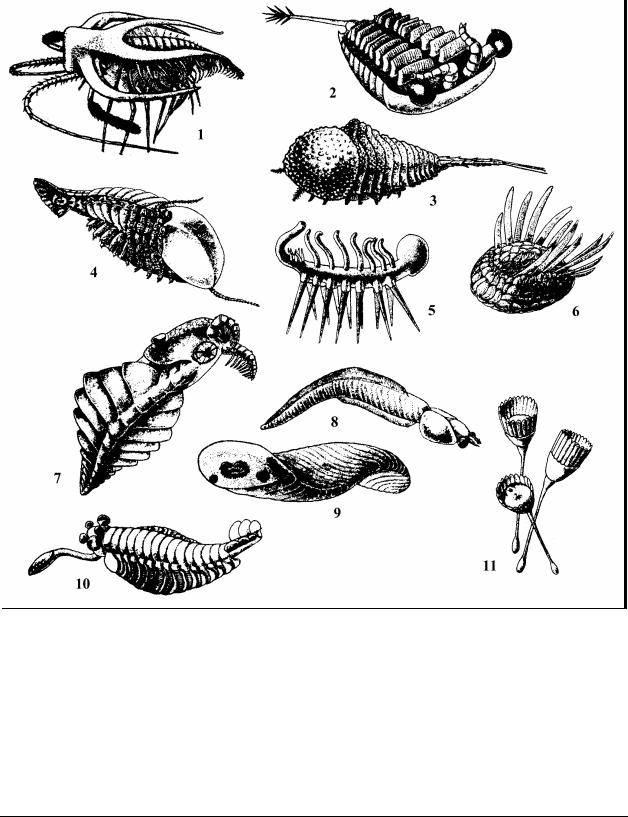
Рис. 9. Реконструкции проблематичных животных кембрийского периода из бургесских сланцев: Тип Arthropoda – Членистоногие, класс не установлен, 1 – Marrella splendens; 2 – Sarotrocercus; 3 – Habelia; 4 – Sidneyia. Проблематичные животные, систематическое положение не известно, 5 – Hallucigenia (3 – 5 см); 6 – Wiwaxia (5 – 6 см); 7 –
Anomalocaris (70 – 100 см); 8 – Nectocaris; 9 – Odontogriphus (около 8 см); 10 – Opabinia; 11 – Dinomischus (4 – 5 см)
39
ОРДОВИКСКИЙ ПЕРИОД (СИСТЕМА)
Длительное время ордовикские отложения рассматривались в составе выделенной ранее силурийской системы. Только в 1879 г английский геолог Ч. Лапворт выделил из силурийской системы нижнюю часть под названием ордовикской системы. Свое название система получила по племени ордовиков, населявших в древности Уэльс (Великобритания). Общепринятых подразделений периода на отделы пока нет. В бывшем СССР ордовик принято было делить на три отдела (Табл. 4). Начало периода около 505 млн., конец – 440 млн. лет назад; общая продолжительность около 65 млн. лет.
В ордовике существовали в основном те же материки, что и в кембрийское время. Лаврентия и Балтика сблизились, от Гондваны отделились Южный и Северный Китай. Для этого периода характерно повышение уровня Мирового океана, что привело к обширной трансгрессии – затоплению морскими водами материковых низменностей. В раннем и среднем ордовике отмечается максимальное за всю фанерозойскую историю Земли распространение эпиконтинентальных морей и резкое сокращение их площадей в позднем ордовике. Сокращение площадей шельфовых морей в конце периода, вероятно, было основной причиной резкого снижения таксономического разнообразия, которое отчетливо проявилось в мелководных морских бассейнах и в меньшей степени в более глубоководных частях шельфов.
Климатические условия ордовика были неоднородны, в этот период можно наметить области сухого жаркого и теплого влажного климата, а также районы возможных похолоданий. По распределению органических остатков морских беспозвоночных можно выделить несколько биогеографических областей.
Таблица 4
Геохронологическое (стратиграфическое) подразделение ордовикской системы
Эпоха (отдел) |
Век (ярус) |
Верхний |
Ашгиллский O3 a |
Средний |
Карадокский O2 k |
|
Лландейльский O2 ll |
|
Лланвирнский O2 l |
Нижний |
Аренигский O1 a |
|
Тремадокский O1 t |
ОРГАНИЧЕСКИЙ МИР
В морях ордовикского периода были широко распространены зеленые водоросли, беспозвоночные и первые позвоночные животные. На суше во второй половине периода появились наземные растения. В морских экосистемах произошло крупнейшее в фанерозое увеличение таксономического
40