

Paleo / MARTYNOV
.pdfли появились на рубеже 1,8 – 1,7 млрд. лет, многоклеточные водоросли на рубеже около 1,0 млрд. лет, но, возможно, и раньше, где-то около 1,4 млрд. лет назад. Таким образом, чрезвычайно длительная эра господства бактерий и близких к ним сине-зеленых водорослей сменилась эрой водорослей, достигавших в водах древних океанов значительного разнообразия форм и красок. Животные появились позднее растений, достоверные одноклеточные известны с рубежа 1,0 млрд. лет, а многоклеточные – около 0,7 млрд. лет назад, хотя не исключено и более раннее появление.
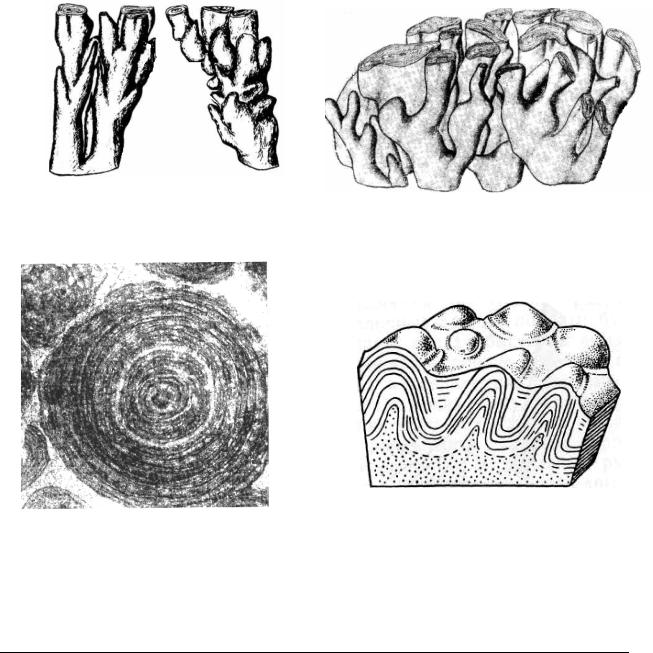
Разнообразие жизни особенно возросло во второй половине протерозоя, в рифее (интервал 1,7 – 0,67 млрд. лет) и в венде (интервал 670 – 570 млн. лет). В конце рифея (700 млн. лет) началась кремнеземная биоминерализация, повидимому, связанная с золотистыми водорослями. Но особенно характерны для рифея и в меньшей степени для венда строматолиты (Stromatolithi), представленные мощными толщами биогенных известняков глобального распространения. Строматолиты были сформированы в результате жизнедеятельности сообществ цианобактерий (Cyanophyta – цианеи), бактерий и других микроорганизмов, явившихся первыми рифостроителями в истории Земли. Древнейшие ископаемые строматолиты были обнаружены в Западной Австралии в породах возрастом 2,8 млрд. лет, хотя проблематичные строматолиты имеют возраст не менее 3,5 млрд. лет. При изучении остатков микроорганизмов, слагающих строматолиты, выяснилась одна интересная особенность, имеющая принципиальное значение. Микрофоссилии разного возраста мало изменяют свою морфологию, что свидетельствуют в целом о консервативности прокариот. Микрофоссилии, относящиеся к прокариотам, оставались практически неизменными довольно длительное время. Во всяком случае, перед нами установленный факт – эволюция прокариот проходила значительно медленнее, чем высших организмов. Строматолиты обитают и в настоящее время, они, как вероятно, и их далекие предки предпочитают теплое мелководье. Строматолитами (от греч. строматос – ковер, литос – камень) – называют тонкослоистые, преимущественно карбонатные постройки. Формы этих построек были чрезвычайно разнообразны: пластообразные, желваковидные, столбчатые и зависели, главным образом, от гидродинамической обстановки и других экологических факторов (Рис. 3). Эти постройки образовывались в результате жизнедеятельности сообществ цианобактерий, бактерий и других микроорганизмов. Выделение извести характерно для бактерий, цианобактерий и различных эукариотических водорослей. Считается, что растения выщепляют СО2 из растворенного в воде бикарбоната кальция. Образующийся при этом кальцит осаждается на слизистых оболочках клеток. Со временем водоросли отмирали и на поверхности, которую занимала колония, оставалась тонкая корочка известкового вещества (Рис. 3: 5). Эта корочка состоит из двух тесно связанных друг с другом слоев, светлого и темного. Предполагают, что светлые, известковые слои формировались весной и летом, когда были благоприятные условия для роста. Осенью и особенно зимой рост водорослей прекращался, выделение извести резко снижалось, и отлагались преимущественно глинистые частицы, оседавшие на поверхность коло-
21

нии из морской воды. К сожалению, наблюдать четкие остатки водорослей в строматолитах удается редко, это связано с тем, что нити цианобактерий не покрывались сплошной известковой корочкой, а отлагали отдельные мелкие зернышки кальцита в слизистом чехле, окружавшем нити. Когда водоросль отмирала, нить распадалась, и зерна кальцита рассыпались по субстрату. За десятки, сотни, а может быть и тысячи лет, такие слои, нарастая друг на друга, образовывали гигантские рифы, поднимавшиеся с морского дна на сотни метров подобно современным коралловым рифам*. Те же водоросли, что образовали строматолиты, или их ближайшие родственники, могут нарастать и на неприкрепленные обломки камней или комочки грунта. Течение и волны перекатывают такие обросшие водорослями желваки, в результате образуются округлые желваковые образования онколиты (Oncolithi) диаметром от нескольких мм до 10 – 12 см – несущие на срезе концентрические круги или радиально лучистые структуры (Рис. 3: 4). Часть желваков, не имеющих правильной формы и внутренней слоистости, называется катаграфии (что означает "древние письмена"). В докембрийских породах онколиты и катаграфии встречаются, как правило, совместно со строматолитами. Водорослевое происхождение большинства этих остатков сомнения не вызывает, хотя не исключено, что часть их образуется при участии животных. Например, часть катаграфий похожа на структуры, образующиеся при активном воздействии на грунт червей илоедов и некоторых роющих организмов.
Для протерозоя характерно изменение в атмосфере баланса метана, углекислого газа и кислорода. Содержание кислорода стало заметно возрастать с уровня 2,2 млрд. лет, что связано с жизнедеятельностью цианобионтов.
Новый этап развития органического мира, начавшийся в среднем рифее, связан с достижением точки Пастера. При таком содержании кислорода в атмосфере (0,01 современного) брожение переходит в окисление (эффект Пастера), начинается формирование озонового щита, что обеспечивало защиту организмов от ультрафиолетового излучения, скрытых только метровым слоем воды.
Животные получили возможность полностью перейти к кислородному дыханию и смогли подниматься к самой поверхности воды. Только переход к кислородному дыханию мог стать важнейшим энергетическим импульсом в развитии многоклеточных организмов. Кислородное дыхание в энергетическом отношении намного выгоднее брожения. Например, при разложении 1 моль глюкозы посредством брожения выделяется энергия в количестве, равном 209,3 Дж, а посредством дыхания – 2872,3 Дж, т.е. в десять раз больше. Именно в позднем рифее, а, возможно, и несколько ранее, произошли события, пока в значительной степени скрытые от нас, результатом которых стал первый экологический взрыв – массовое появление и расцвет вендской флоры и фауны.
* Применяемые ранее к строматолитам бинарные названия не соответствуют требованиям кодекса номенклатуры, и для них предложено не вводить латинских названий, обозначая различные формы буквенными символами. К настоящему времени предложено более 20 классификаций строматолитов.
22
1 |
2 |
3
5
4
Рис. 3. Известковые фоссилии водорослевого происхождения: 1, 3 – Gymnosolen ramsayi – верхнерифейские строматолиты; 2 – Linella avis – строматолиты вендского комплекса; 4 – онколиты из группы озагия – Osagia columnata – средний рифей (поперечный срез); 5 – схема продольного среза через строматолит
23
ВЕНДСКИЙ ПЕРИОД (СИСТЕМА)
(венды, венеды – название древнего славянского племени, населявшего север и запад России)
Венд как переходные слои от протерозоя к кембрию был выделен в 1952 г Б.С. Соколовым на территории Прибалтики, здесь же находится и стратотип венда. Начало венда датируется примерно 670 млн. лет назад, продолжительность около 80 – 100 млн. лет. Ввиду уникальности биоты этого времени и независимости всей вендо-эдиакарской биоты бесскелетных организмов как от предшествующих биот позднего рифея, так и от последующих кембрия, по отношению к которым ее даже нельзя считать переходной, обособлен в вендский период.
ОРГАНИЧЕСКИЙ МИР
Палеонтологическая характеристика венда по своему разнообразию и уровню развития животных и растительных организмов резко отличается от предшествовавшего периода, хотя на уровне "микробиот" и биолитов (строматолиты), несомненно, существует преемственность. Для венда характерен первый максимум биоразнообразия. В венде начался третий важнейший этап развития органического мира докембрия – этап становления основных типов животного мира, и прежде всего, многоклеточных. Вендская фауна и флора отличались большим разнообразием и богатством форм, имеющих планетарное распространение. Флора в венде была представлена многоклеточными водорослями – метафитами и разнообразными одноклеточными водорослями (в том числе цианофитами). Находки микрофоссилий, внешне сходных со спорами высших растений, в докембрии и нижнем палеозое порождали гипотезы, допускавшие заселение суши задолго до появления в геологической летописи первых остатков достоверных наземных растений. Обнаруженные в верхах докембрия споры с отчетливой трехлучевой складкой, принадлежали, скорее всего, к проблематическим водорослям (Vendotaenides), встречающимся в тех же слоях, а не высшим растениям.
Сообщество вендских животных состояло из бесскелетных организмов: кишечнополостных (медуз, полипов, морских перьев); организмов, близких к червям и членистоногим; сабеллидит (Sabellidites), проблематичных иглокожих и конодонтов (Рис. 4). Отпечатки этих животных впервые найдены в 1947 г Р. Сприггом в Южной Австралии, в районе рудника Эдиакара (к северу от Аделаиды), в связи с чем эту фауну часто называют "эдиакарская фауна", хотя в дальнейшем она была обнаружена в вендских отложениях европейской части России, Украины, Англии, Канады, Африки и других мест.
Отличительной особенностью животных вендских морей было отсутствие у подавляющего большинства минерального скелета. Это были мягкотелые животные. Однако покровы их тела достигали значительного уплотнения, благодаря чему на мягком илистом грунте при жизни или после захоро-
24
нения оставались четкие отпечатки.
Основу вендской фауны составляли кишечнополостные – 67%, около 25% – черви и около 5% – членистоногие. Следует отметить, что оценка систематической принадлежности большинства вендских организмов крайне спорна и условна, поскольку всегда остается вероятность наличия жизненных форм, не имеющих аналогов в современной фауне.
Кишечнополостные были наиболее крупными животными в вендской фауне, достигавшие длинны до 1 м, и в подавляющем большинстве обладавшие радиальной симметрией (Radialia). Это были медузоподобные организ-
мы, вероятно, парившие в толще воды (Ediacara flindersi, Beltanella gilesi,
Medusinites asteroides и др.). В составе вендской фауны были и двустороннесимметричные животные (Bilateria), однако по сравнению с радиальносимметричными они были менее разнообразны. Среди этих животных были несегментированные и сегментированные формы. Несегментированные (Protechiurus) – наиболее примитивные, листовидные животные длиной до 70 мм, условно относимые к плоским червям. Подавляющее большинство вендских билатерий является сегментированными организмами, например гигантские Diskinsonia, достигающие длины 1 м при толщине тела 3 мм. Предполагают, что эти организмы образуют отдельный тип. Своеобразную группу в вендской фауне составляли мелкие сегментированные животные, интерпретируемые как возможные предки членистоногих и сочетавшие в своем строении признаки примитивных членистоногих и червей-полихет (Praecambridium sigillum). Внешне они сходны с личинками трилобитов.
В состав вендской фауны входили и сабеллидиты (Sabbelitida) – представители одной из немногих групп животного мира, которые продолжали существовать в фанерозое. Их скелеты сохранились в виде одиночных, тонких и длинных трубок, состоящих из хитиноподобного вещества. Среди вендских организмов встречено значительное количество колониальных перистовидных форм, похожих на современных альционарий, или морские перья (Charniodiscus). Колониальные формы отличались крупными размерами и простым строением.
Детальное изучение вендской фауны многоклеточных выявило ее специфические особенности, главными из которых являются: отсутствие или слабое развитие скелетных элементов, большое разнообразие жизненных форм, резкое преобладание кишечнополостных, присутствие всех основных экологических групп организмов и др. Вендская фауна в целом характеризуется высокой морфологической дифференциацией, отражающей уже сложившиеся таксоны очень высокого ранга, до типов включительно. Видовое разнообразие на этом уровне дифференциации довольно незначительно. Надежные данные о дивергенции таксонов высокого ранга в венде пока отсутствуют, все типы и классы вендской фауны появляются как бы внезапно. Для объяснения этого явления приходится допустить необычайно быстрое формирование всех основных типов беспозвоночных в геологически очень короткое время.
25

1а |
1б |
2а |
2б 3
4 |
5 |
6 |
7 |
10
8 |
9 |
Рис. 4. Бесскелетная фауна венда: 1 – Nemiana (а – реконструкция, б – аборальная сторона), 2 – Cyclomedusa (а – реконструкция, б – аборальная сторона), 3 – Staurinidia - (план строения), 4 – Cyclomedusa, 5 – несегментированное двусторонне-симметричное животное Protechiurus, 6 – Albumares – форма с трехлучевой симметрией, 7 – кольчатый червь Diskinsonia, 8 – Praecambridium – загадочное животное, возможно предок моллюсков, 9 – Vendia sokolovi – возможный предок трилобитов, 10 – Charniodiscus – перистовидный колониальный полип.
26
Это предположение можно косвенно подтвердить следующим наблюдением: 60 из 90 классов морских многоклеточных появились в вендокембрийском интервале, а от перми до наших дней (примерно за 280 млн. лет) – только пять. Кроме того, вендская фауна, по своему составу, имеет мало общего как с предшествующей ей фауной рифея, так и с последующей скелетной фауной кембрия, по отношению к которым она не может рассматриваться даже как переходная. Ни одна группа организмов венда не дала потомков (кроме сабеллидит – рассматриваются рядом авторов как предки погонофор, но в тоже время ультраструктура трубок погонофор и сабеллидит по мнению А. Урбанека принципиально несопоставимы). Прямые связи вендской и кембрийской фауны не установлены, возможно, они принадлежат к независимым линиям развития. Сходство вендских фаунистических ассоциаций в разных регионах мира, отражающее отсутствие существенных экологических барьеров, дало возможность использовать биостратиграфические методы для расчленения и корреляции отложений венда.
Ф А Н Е Р О З О Й С К И Й Э О Н
(греч. phaneros – видимый, явный; zoe – жизнь)
Фанерозойский эон начался около 600 – 530 млн. лет назад и продолжается до настоящего времени. Фанерозой палеонтологически документирован несравнимо полнее криптозоя. Граница между фанерозоем и криптозоем проводится по появлению у самых разнообразных организмов, как ранее известных, так и вновь возникших, способности строить минеральный скелет. Способность к скелетообразованию носила массовый характер и в геологически короткое время проявилась почти у всех царств и типов животных. Фанерозой принято разделять на три эры: палеозойскую, мезозойскую и кайнозойскую. Палеозойская эра (наиболее продолжительная 350 млн. лет) может быть разделена на две части (подэры) – ранний и поздний палеозой (метазой), иногда им придается ранг самостоятельных эр.
ИЗМЕНЕНИЕ ЖИВОТНОГО МИРА ПЛАНЕТЫ НА НИЖНЕЙ ГРАНИЦЕ КЕМБРИЯ
На нижней границе кембрия происходят наиболее значительные изменения в характере органического мира, послужившие основой для разделения всей истории Земли на криптозой и фанерозой. На этом рубеже появляется огромное количество скелетных остатков, принадлежащих животным практически всех современных типов. Примечателен и тот факт, что скелетные образования появляются практически "одновременно" у организмов, находящихся на разных уровнях эволюционного развития, от раковин моллюсков до панцирей членистоногих.
В то же время превосходная сохранность отпечатков бесскелетных может свидетельствовать о том, что в венде происходило прогрессивное уплотнение тканей, связанное с их совершенствованием и дифференциацией. Как
27
продолжение этого процесса можно рассматривать активную склеротизацию тканей и массовое появление в кембрии скелетной фауны. Многие специалисты высказывали сомнения по поводу возможности одновременного появления скелетных образований у разных систематических групп и надуманности данной проблемы. Однако самые тщательные проверки показали отсутствие скелетных образований в докембрии. Выдвигалось предположение о том, что скелетные остатки в докембрии имели микроскопические размеры. Однако данное утверждение не находит подтверждений, в настоящее время хорошо разработаны и активно применяются методики извлечения микрофоссилий, но микроскелетных остатков так и не обнаружено.
Для объяснения причин этого события выдвинут целый ряд разнообразных гипотез, которые в общем можно разделить на несколько основных групп.
1.Сторонники химического, геохимического и биохимического направления считают, что химизм вод докембрийских морей просто не давал возможности организмам строить твердые скелеты. Отсутствие скелета у животных позднего протерозоя, возможно, было вызвано высоким содержанием углекислоты в атмосфере и гидросфере. Это приводило к резкому увеличению растворимости извести и затрудняло ее извлечение из воды. Подчиняясь чисто химическим законам, раковины просто не могли возникнуть. А если и возникали, то тут же растворялись. Данные гипотезы не могут быть приняты на том основании, что в этот же период формировались мощнейшие толщи карбонатных пород, а водоросли строили мощные рифоподобные постройки-биогермы. В настоящее время при постройке скелета животными используется около 40 различных минералов, такой широкий выбор позволяет всегда подобрать альтернативные материалы. Кроме того, в начале кембрия у животных появились не только карбонатные, но и кремневые, фосфатные скелеты.
2.Многие биологи на основании того, что нигде в мире не известно ни одного поврежденного остатка докембрийских бесскелетных животных объясняют это отсутствием хищников. В этот период, по-видимому, не существовало активных хищников, а также организмов, питавшихся крупными пищевыми частицами (в том числе и трупоедов). Таким образом, появление скелета – результат активизации (появления) хищников. Предположение достаточно сомнительно хотя бы на том основании, что и в современных морях количество бесскелетных организмов огромно, несмотря на активность хищников.
Неправдоподобным представляется и предположение о необходимости утяжеления организмов в связи с переходом к жизни на морском дне как элемент противодействия смыву течениями. Предположение не логично, поскольку скелеты самой различной природы имеют не только бентосные формы, но и планктон.
Оригинальную гипотезу выдвинули американские исследователи Д. Ламар и П. Мерифельд, которые связали появление скелетов с образованием Луны. В морях возникли приливы и отливы, что значительно усилило
28
динамику прибрежных вод, и скелеты стали абсолютно необходимы. Эта гипотеза отпала сама собой после определения возраста лунных пород.
3.В третью группу можно выделить "мутационные" гипотезы. В данном случае одновременное появление скелетов связывают с какими-то внешними явлениями. Например, немецкий палеонтолог О. Шиндевольф связывает подобные скачки в развитии животного мира с резким увеличением уровня космической радиации. В соответствии с его гипотезой, по соседству с Солнечной системой вспыхнула сверхновая звезда, и возросший поток космических лучей обрушился на Землю, вызывая массовые мутации у самых различных организмов. Как известно, подавляющее большинство мутаций снижает жизнеспособность мутантных особей, чем и пытаются объяснить массовое вымирание видов прежней фауны. Общее усиление мутационного процесса, по мнению Шиндевольфа, приводит к возникновению и быстрому распространению новых форм. При этом возникают новые типы организации. В то же время, мутации – сложно предсказуемое и ненаправленное явление, в связи с чем сложно себе представить, что организмы, находящиеся на разных уровнях организации, отреагировали одинаково – образованием скелетов. И наконец, расчеты ряда авторов показали, что вспышки Сверхновых звезд не могут вызвать такого повышения радиационного фона на поверхности Земли, которое имело бы предполагаемый Шиндевольфом мутагенный эффект.
4.Целый ряд теорий связывают происхождение скелета с эволюцией атмосферы Земли. Большой интерес в решении этой проблемы представляет гипотеза Л. Беркнера и Л. Маршалла. В центре внимания этих авторов стоит анализ зависимости развития жизни на Земле от содержания кислорода в Земной атмосфере, медленно, но неуклонно повышавшегося в ходе развития жизни. Одним из важнейших, переломных моментов в развитии жизни было достижение точки Пастера. Достижение пастеровской точки в развитии атмосферы Земли ознаменовалось не только появлением возможности аэробной диссимиляции организмов. При содержании кислорода в атмосфере в количестве 0,01 от современного, формирующийся озоновый экран может защитить от жесткой ультрафиолетовой радиации уже и верхние слои воды (требуется только "помощь" слоя воды толщиной примерно 1 м). Это позволило не только подниматься в поверхностные слои водоемов, наиболее богатые солнечной энергией, но и резко усилить эффективность фотосинтеза, а следовательно, увеличить биопродуктивность и выделение свободного кислорода. Как следствие резко расширяется арена жизни: ставшие доступными малые глубины водоемов предоставляют более разнообразные условия обитания, чем средние и большие глубины. Освоение новой, открывшейся арены неминуемо должно было привести к резкому повышению разнообразия форм жизни и взрыву формообразования.
Несколько иначе подходит к данной проблеме К. Тоув, в основе его гипотезы также лежит концентрация кислорода. По его мнению, у докембрийских животных в условиях малого содержания кислорода в окружающей среде не могло быть хорошо развитых соединительнотканных образований,
29
формирующих основу для развития скелета. Прочность соединительнотканных образований базируется, прежде всего, на содержании в них белка коллагена, в состав которого входит аминокислота оксипролин. Реакция образования оксипролина возможна только при наличии достаточно большого количества кислорода в окружающей среде. При низком содержании кислорода синтез коллагена был биохимически затруднен, и организмы не могли иметь прочных соединительнотканных образований, а следовательно и скелетов. Слабой стороной этих гипотез является отсутствие убедительных доказательств резкого отличия условий в венде и кембрии. Кроме того, существуют не менее убедительные доказательства существования атмосферы близкой по составу к современной, по крайней мере, 3 млрд. лет назад.
5. Одной из особенностей границы кембрия и докембрия является активное фосфоритообразование. Именно к этому периоду приурочено образование целого ряда крупных месторождений фосфоритов. Возможно, кембрийская мутация связана с массовым поступлением в морские воды огромного количества вулканогенного фосфора.
Но данному факту можно дать и обратное толкование. Поскольку фосфор
– один из обязательных элементов всех живых существ, то возможно массовое отложение фосфоритов на границе кембрия и докембрия указывает на катастрофическое вымирание вендских организмов. Факт массового вымирания в конце венда подтверждается палеонтологическими данными, по крайней мере, наиболее характерные элементы вендо-эдиакарской фауны не встречаются в кембрии.
Таким образом, у нас пока нет явных свидетельств экологического характера, объясняющих отсутствие тканевой минерализации в докембрии. Многочисленные гипотезы, высказанные по этому поводу, не выдерживают критики. Кроме того мы не можем указать существенных различий в обстановке венда и кембрия, скорее они были сходны.
Нельзя не отметить и тот факт, что многие группы организмов, характерных для венда, не обладали скелетом в силу своей организации, так же как и сейчас не имеют скелета медузы, плоские черви, многие полипы, аннелиды. В то же время первые этапы склеротизации, по всей видимости, проходили уже в венде, на что могут косвенно указывать резко рельефные отпечатки некоторых вендских форм (птеридинид), что позволяет предположить наличие у них достаточно жестких, вероятнее всего, органических скелетных элементов.
30