

Цикл пуриновых нуклеотидов

Выделяющийся аммиак должен элиминировать в виде глутамина,аспарагина илиаланина.
Глу + NH3+ АТФ → Глн + АДФ + Фн (глутамин-синтетаза)
Асп + Глн → Асн + Глу (аспарагин-синтетаза)
ПВК + Глу ↔ Ала + a-кетоглутарат (аланин-аминотрансфераза)
Эти аминокислоты с кровотоком достигают печени, где используются как субстраты ГНГ. Т. обр., мышцы играют важную роль в межорганном обмене субстратами (циклы Кори и Фелига).
Пути утилизации атф в мышце
Механическая работа – мышечное сокращение.
Работа АТФ-аз, обеспечивающая электрическую составляющую:
Na+/K+-АТФ-аза – способствует поддержанию потенциала покоя; транспорт субстратов.
Ca2+-АТФ-аза – мышечное расслабление,
Mg2+-АТФ-аза – стабилизация АТФ, и т.д.
Расслабление мышц– АТФ-зависимый процесс (откачиваниеCa2+из саркоплазмы).
Благодаря мышцам химическая энергия превращается в механическую
Мышцы являются самой значительной, по массе, тканью тела человека. Сразу после рождения она составляет 25%, в юношеском возрасте - 40% и у пожилых людей - немногим менее 30% массы тела.
Эффективность превращения химической энергии в механическую при сокращении мышц зависит от ряда факторов. (1) Необходим постоянный приток химической энергии. У позвоночных эту функцию выполняют АТФ и креатинфосфат. (2) Необходимы средства регуляции механической активности, в частности, скорости, длительности и силы сокращения (если речь идет о мышце). (3) Сократительный аппарат должен быть соединен с оператором (в биологических системах роль оператора выполняет нервная система). (4) Механизм превращения химической энергии в механическую должен иметь возможность возвращения в исходное состояние.

Рис.17.2.Поперечно-полосатая мышечная ткань, вид под микроскопом
Всем этим требованиям в организме отвечают три типа мышц: скелетные мышцы, сердечная мышца и гладкие мышцы. Скелетная и сердечная мышцы под микроскопом имеют поперечно-полосатое строение (рис.17.2). В гладких мышцах такой исчерченности нет. С другой стороны, сокращение поперечно-полосатых мышц контролирует нервная система. Оно происходит в результате проведения нервного импульса. Для сокращения сердечной и гладких мышц специального стимулирующего действия нервной системы не требуется. Через те нервы автономных ветвей нервной системы, которые идут к этим мышцам, регулируется лишь сила и длительность сокращений.
В саркоплазме мышечных клеток находятся АТФ, креатин фосфат и ферменты гликолиза
Поперечно-полосатые мышцы состоят из волокон, которые являются многоядерными мышечными клетками, окруженными электрически возбудимой мембраной - сарколеммой. Такая клетка может простираться на всю длину мышцы. Она содержит в своем составе пучок большого количества параллельно расположенных миофибрил, погруженных во внутриклеточную жидкость - саркоплазму. В саркоплазме содержатся гликоген, макроэргическое соединение АТФ, креатин фосфат и ферменты гликолиза.

Рис.17.3.Поперечно-полосатая мышца в поперечном сечении
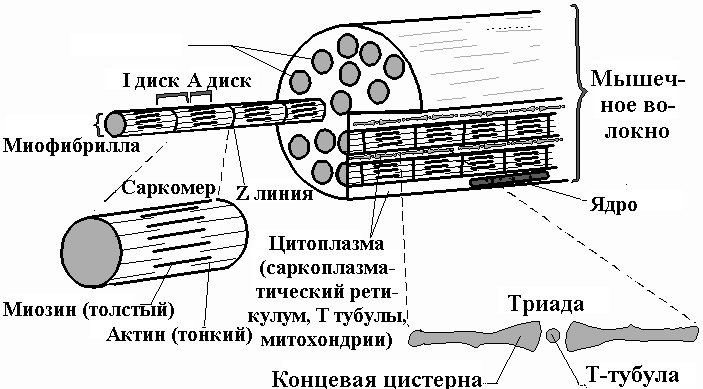
Рис.17. 4.Схематическое строение мышечного волокна
Функциональной единицей мышцы является саркомер
![]()
![]()

![]()
![]() Рис.
17.5.Структура саркомера
Рис.
17.5.Структура саркомера
По оси мышечного волокна расположены повторяющиеся участки размером 1500-2300 нм (рис.17.4) - саркомеры. При рассмотрении миофибриллы под электронным микроскопом видны перемежающиеся темные и светлые полосы или диски (А и Iдиски). Центральная зона А диска (Hзона) при этом кажется менее оптически плотной, чем остальная его часть.Iучасток (илиIдиск) как бы рассекается на две части очень плотной и узкойZ-линией (рис.17.5).
Поперечная исчерченность скелетных и сердечной мышц, видная уже под световым микроскопом (рис.17.2), обусловлена высокой степенью организации, при которой большинство мышечных клеток-волокон располагаются в ряд таким образом, что их саркомеры (из разных волокон) строго параллельны друг другу.
В толстых нитях находится миозин, в тонких - актин, тропомиозин и тропонин
При рассмотрении миофибриллы под электронным микроскопом видны два типа вытянутых нитей. Один тип - это толстаянить, соответствующая А диску. В ней преимущественно содержится белок-миозин. Толстые нити имеют в диаметре 16 нм и на поперечном разрезе уложены в форме шестиугольника (рис.17.6).

Рис. 17.6.Поперечное сечение через разные части миофибриллы
Другой тип - тонкая
нить, расположена в I![]() диске и проходит в А диск, не достигая
Н зоны в А диске (рис.17.5, 17.7). В тонких
нитях содержится белок-актин, тропомиозин
и тропонин (рис. 17.10). В А диске тонкая
нить накручена на толстую (миозин) таким
образом, что на поперечном разрезе
такая структура также имеет гексагональную
форму (рис.17.4). Каждая тонкая нить лежит
симметрично между 3 толстыми нитями, а
каждая толстая нить симметрично окружена
6 тонкими нитями.
диске и проходит в А диск, не достигая
Н зоны в А диске (рис.17.5, 17.7). В тонких
нитях содержится белок-актин, тропомиозин
и тропонин (рис. 17.10). В А диске тонкая
нить накручена на толстую (миозин) таким
образом, что на поперечном разрезе
такая структура также имеет гексагональную
форму (рис.17.4). Каждая тонкая нить лежит
симметрично между 3 толстыми нитями, а
каждая толстая нить симметрично окружена
6 тонкими нитями.
Взаимодействие толстых и тонких нитей осуществляется с помощью поперечных мостиков, которые представляют собой домены молекул миозина. Они выявляются с интервалом 14 нм вдоль толстых нитей и перекрывают промежуток шириной 130Å между поверхностью толстых и тонких нитей (рис. 17.7). По существу, именно взаимодействие миозиновых поперечных мостиков с единицами актина в тонких нитях генерирует силу сокращения.


Рис. 17.7.Схематическое изображение структуры толстых и тонких нитей и их взаимного перекрывания. На толстых нитях головки миозина образуют поперечные мостики