

Gistologia_v_voprosakh_i_otvetakh_Sluka
.pdfконтактирующих клеток сближаются на расстояние в 2-3 нм и с помощью интегральных белков коннексонов они формируют тубулы, имеющие поры. Тубулы соседних клеток соединены друг с другом, поэтому диффузия веществ между двумя клетками идет без выхода их в межклеточное пространство. Так построены щелевидные соединения (нексусы) — между кардиомиоцитами и гладкими миоцитами. Нексусы позволяют передавать возбуждение с одной клетки на другую.
Кэтойгруппеконтактовотносятсяихимические синапсы— контакты между нейронами или нейроном и мышечным волокном. Синапсы осуществляют одностороннюю передачу возбуждения или торможения.
7.Цитоплазма. Общаяморфофункциональнаяхарактеристика. Гиалоплазма. Цитоскелет: организация и функциональное значение
Цитоплазма клетки включает в себя гиалоплазму и находящиеся в ней обязательные компоненты органеллы, а также различные непостоянные структуры — включения.
Гиалоплазма (цитозоль) — матрикс цитоплазмы — сложная коллоидная система, включающая в себя биополимеры: белки, нуклеиновые кислоты, полисахариды. В гиалоплазме содержатся ферменты метаболизма Сахаров, аминокислот, липидов, азотистых оснований. Система способна переходить из золеобразного (жидкого) состояния в гелеобразное и обратно. При этом изменяется скорость реакций и перемещения веществ по цитоплазме. В гиалоплазме с участием рибосом идет синтез белков для клетки. В гиалоплазме находятся внутриклеточные рецепторы. Цитоскелет — это опорный каркас клетки, который обеспечивает ей сохранение формы, создает условия для перемещения но цитоплазме органелл и различных веществ, а также перемещение самой клетки и осуществление ею экзо- и эндоцитоза.
Строение цитоскелета. Компонентами цитоскелета являются: микротрубочки, микрофиламенты и промежуточные филаменты.
Микротрубочки — непостоянные удлиненные структуры клетки, имеющие цилиндрическую форму и образованные 13-ю филаментами тубулиновой природы. Толщина микротрубочки составляет 25 нм и диаметр канала — 15 нм. Они могут расти. В цитоплазме существует равновесие между микротрубочками и растворенным тубулином: с одного конца трубочки распадаются, с другого — вновь образуются. Не распадаются микротрубочки центриолей, базальных телец, ресничек и жгутиков.
21
При митозе микротрубочки цитоскелета распадаются, а из освободившегося тубулина образуется веретено деления. После митоза процесс идет в обратном направлении. Если клетку обработать колхицином, разрушающим микротрубочки, клетки теряют способность делиться, изменяется их форма.
Микрофиламенты — опорные, фибриллярной природы структуры, входящие в состав субмембранного опорно-сократитель- ного аппарата клетки. Существует три типа филаментов:
•микрофиламенты толщиной 5 6 нм (актиновые);
•толщиной 10 нм (миозиновые);
•толщиной около 7 нм (промежуточные).
Актиновые и миозиновые филаменты образуют миофибрил-
лы в миоцитах и мышечных волокнах, в других клетках обеспечивают сокращения и перемещение клетки, процессы эндо- и экзоцитоза, формирование псевдоподий и микроворсинок. Эти филаменты участвуют в сокращении тромбов. Много филаментов содержится в подмембранном слое клеток. С ними связаны интегральные белки мембран.
Промежуточные филаменты — наиболее стабильные филаменты. Главная их функция — опорная. Они выявляются по всей длине отростков нервных клеток, в цитоплазме миоцитов, в десмосомах и т. д., имеют тканевую специфичность и в разных тканях построеныизразныхбелков. Вэпителииониимеюткератиновуюприроду (тонофибриллы), в клетках мезенхимного происхождения — состоятизвиментина, вмышцах— издесминаискелетинаит. д.
Клеточный центр — клеточная органелла немембранной природы, обеспечивающая организацию микротрубочек, их сборку из тубулиновыхсубъединиц. Состоитизцентриолейицентросферы.
Центриоли — парные образования цилиндрической формы, расположенные перпендикулярно друг другу и состоящие из микротрубочек. Стенку центриоли образуют 9 триплетов микротрубочек А, В и С, расположенных по окружности и связанных белковыми мостиками — динеиновыми "ручками". В центральной части центриоли микротрубочки отсутствуют. Триплеты центриоли связаны с сателлитами — сферическими тельцами диаметром 75 нм. Сателлиты являются местом сборки микротрубочек.
Центросфера — совокупность микротрубочек, отходящих от сателлитов центриолей. Центриоли являются не только центром сборки микротрубочек для цитоскелета, но и для образования некоторых органелл специального назначения — ресничек и жгутиков. Кроме того, центриоли являются одним из центров формирования ахроматинового веретена деления при митозе.
22
8.Органеллыцитоплазмы. Классификация органелл, ихфункции. Структурнаяорганизация
синтетическогоаппаратаклетки: эндоплазматическая сеть, рибосомы, комплекс Гольджи
Органеллы цитоплазмы — это постоянно присутствующие и обязательные для клеток микроструктуры цитоплазмы, специализированные на выполнении жизненно важных функций.
Классификация органелл. Существует две классификации. По одной из них различают мембранные и немембранные органеллы. По второй они подразделяются на органеллы общего назначения (постоянно присутствующие и обязательные для всех клеток); и органеллы специального назначения (реснички, жгутики, микроворсинки, тонофибриллы, миофибриллы), характерные для специализированных клеток. Органеллы специального назначения, как правило, являются производными органелл общего назначения.
Мембранные органеллы представляют собой одиночные или связанные друг с другом отсеки цитоплазмы, отграниченные мембраной от окружающей их гиалоплазмы. К ним относятся: митохондрии, эндоплазматическая сеть, аппарат Гольджи, лизосомы, пероксисомы.
Немембранные: свободные рибосомы и полисомы, клеточный центр, микротрубочки и микрофиламенты цитоскелета, микроворсинки, реснички и жгутики.
Свободные рибосомыиполирибосомы— этомелкие, округлые,
немембранные органеллы, состоящие из большой и малой субъединиц. Каждая субъединица содержит молекулу рибосомальной РНК и специальные белки, которые синтезируются в цитоплазме, азатемпоступаютвядрышко, гдесоединяютсясрРНК. Субъединицы рибосом выходят в цитоплазму. Полирибосомы — группа рибосом, связанных молекулой информационной РНК.
Рибосомы, лежащие в цитоплазме поодиночке или группами — это отдельный класс органелл. Они участвуют в синтезе белков цитоплазмы (цитозоля и органелл) и ядра, обеспечивая возобновление разрушающихся белков и потребности роста, создание особых белков (гемоглобин).
Свободные рибосомы характерны для недифференцированных юк (цитоплазма базофильна). Обычно белок синтезируется не на одной, а на группе рибосом, связанных малыми субъединицами С молекулой информационной РНК. Аминокислоты к рибосоме переносятся транспортной РНК. Рибосома создает условия для взаимодействиямеждутранспортнойиинформационнойРНКиобеспечиваетсозданиеполипептидныхсвязеймеждуаминокислотами.
23
Белки, синтезируемые на свободных полисомах, используются клеткой для себя, а на связанных с мембранами эндоплазматической сети — в основном на экспорт путем секреции.
Эндоплазматическая сеть — это мембранная органелла общего назначения, представленная цистернами, мешочками, трубочками, которые в совокупности создают в цитоплазме целостную систему, предназначенную для синтеза белков, липидов и углеводов, а также внутриклеточного транспорта веществ. Раз-
личают гранулярную (rЭПС) и агранулярную (аЭПС) цитоплаз-
матическую сети; они связаны между собой переходной сетью. В гранулярной сети мембранные цистерны со стороны гиалоплазмы покрыты рибосомами и полисомами. В мембране гЭПС находятся белки, обеспечивающие прикрепление и связывание рибосом, а также уплощение цистерн. В гЭПС по программе информационной РНК (и-РНК), с которой связаны рибосомы, создается полипептидная цепь из приносимых с помощью транспортной (т-РНК) аминокислот. Начальный конец полипептидной цепи — "сигнальный участок", прикрепляется к мембране, а затем проходит через нее внутрь цистерны. Здесь он отрезается с помощью ферментов, а молекула белка подвергается конформации. В дальнейшем белок транспортируется в комплекс Гольджи, а оттуда, в виде окруженных мембраной пузырьков — за пределы клетки (на экспорт). Этим же способом созда-
ются белки лизосом и интегральные белки мембран. Гранулярная эндоплазматическая сеть сильно развита в клет-
ках, которые активно синтезируют белки (фибробласты, плазматические клетки, гепатоциты и др.). Для белоксинтезирующих клеток характерным признаком является базофилия цитоплазмы.
Агранулярная сеть представлена мембранными трубочками, пузырьками и цистернами, не содержащими рибосом; образуется из гранулярной сети, которая теряет рибосомы. В клетках аЭПС по объему меньше, чем rЭПС. Она принимает участие в завершающих этапах синтеза липидов (холестерин), некоторых полисахаридов (гликоген), накапливает соли Са2+. Последняя функция связана с тем, что в мембранах некоторых клеток (мышечные) содержатся кальциевые каналы, кальциевые насосы и кальций связывающие белки.
Агранулярная сеть хорошо развита в клетках, которые синтезируют стероидные гормоны (интерстициальные клетки половых желез, клетки коры надпочечников) и принимающих участие в обезвреживании токсинов (клетки печени).
Комплекс Гольджи (пластинчатый комплекс) — структурно и функциональнополяризованнаямембраннаяорганелла, состоящая
24
из плоских цистерн или мешочков, которые наподобие блюдечек сложены в стопки, а также транспортных пузырьков и вакуолей, конденсирующих секрет гранул. Таких зон, названных диктиосомами, в клетке может быть несколько. Они функционально и структурно связаны между собой. В комплексе Гольджи различают проксимальный или формируемый, обращенный к ЭПС цис-полюс, и дистальный или сформированный, обращенный к плазмолемме транс-полюс.
Функции. В комплексе Гольджи осуществляется упаковка, конденсация и выведение белковых секретов, синтез углеводов и присоединение их к полипептидам, формирование лизосом. Гранулы, наполненные секретом, непрерывно отделяются со зрелой поверхности диктиосомы, подходят к плазмолемме и выделяют содержимое за пределы клетки Из гранулярной эндоплазматической сети продукты синтеза переносятся к цисповерхности комплекса Гольджи транспортными пузырьками, при этом, транспорт белков происходит в трех направлениях — в гидролазные пузырьки (первичныелизосомы), вплазмолемму ивсекреторные гранулы.
9.Аппаратвнутриклеточногоперевариванияизащиты: эндосомы, лизосомы, пероксисомы. Структурная организация, функции. Рольгетерофагиииаутофагиив нормальнойжизнедеятельностиклеток
Аппарат внутриклеточного переваривания — это совокупность мембранных везикул с кислым содержимым (эндосомы) и лизосом, которые предназначены для обеспечения катаболических процессов в цитоплазме клетки. Они объединены в единую систему в связи с наличием в мембранах АТФ-зависимого протонного насоса, создающего низкие значения рН в цитоплазме клеток.
Процесс внутриклеточного переваривания осуществляется последовательно— ранняяэндосома-поздняяэндосома-лизосома.
Эндосомы — мембранные пузырьки с постепенно закисляющимся содержимым, обеспечивающие частичный или полный гидролизмакромолекул, предшествующийлизосомальномугидролизу.
Ранние эндосомы — мембранные пузырьки, расположенные по периферии цитоплазмы, в которых при наличии слабокислой среды с помощью протеаз происходит частичное переваривание макромолекул — отщепление лигандов от рецепторов, с возможным возвращением последних назад в плазмолемму.
Поздние эндосомы — образуются позже предыдущих и лока- ш чуются в зоне ядра. Их внутренняя среда более кислая, и матрикс является электронно-плотным. Ферменты поздних эндосом
25
осуществляют более сильный уровень переваривания (расщепляются лиганды). Оставшиеся ферменты и продукты частичного гидролиза затем направляются в лизосомы.
Лизосомы — это шарообразные тельца, имеющие 0,2-0,4 мкм в диаметре, ограниченные мембраной, содержащие гидролитические ферменты. Основной из них — кислая фосфатаза. Ферменты расщепляют биополимеры.
Различают первичные лизосомы (гидролазные пузырьки) и вторичные— фаголизосомы, аутофаголизосомы, остаточные тельца.
Первичные лизосомы или гидролазные пузырьки содержат фер-
менты в неактивной форме. Предполагается, что гидролазные пузырьки обеспечивают транспорт протеолитических ферментов из комплекса Гольджи. Их перемещение происходит с участием микротрубочек цитоскелета.
Вторичные лизосомы — это мембранные органеллы диаметром 0,5-2 мкм, формирующиеся с участием поздних эндосом и осуществляющие конечные этапы внутриклеточного гидролиза макромолекул. При контакте с субстратом ферменты лизосом разрушают его до мономеров, которые могут утилизироваться клеткой. Начальный процесс слияния поздней эндосомы или лизосомы с субстратом, подлежащим перевариванию, называют
фаголизосомой или гетерофаголизосомой. Если объектом гидролиза являются химические компоненты самой клетки, то образующуюся структуру называют аутофаголизосомой. Если разрушение идет не до конца и в фаголизосоме остаются непереваренныепродукты, онапревращаетсявостаточноетельце.
Роль гетеро- и аутофагии. Гетерофагия осуществляет важ-
ную роль в жизнедеятельности клеток, обеспечивая нормальный гидролиз различных веществ. В случаях недостаточной активности лизосомальных ферментов, их дефиците, в клетках происходит накопление не переваренных остатков различных веществ (болезни накопления).
Роль аутофагии в жизнедеятельности клеток состоит в по-
стоянном обновлении внутриклеточных структур, подвергающихся изнашиванию. Происходит постоянное переваривание фрагментов клеточных органелл и мембран с заменой их новыми структурами. Предполагается, что аутофагия осуществляет отбор и уничтожение дефектных структур, а также может быть реакцией на недостаточность трофики клетки.
Пероксисомы — мембранные органеллы, образованные пузырьками с диаметром 0,2-1,5 мкм, содержащими однородный матрикс средней электронной плотности и уплотненную сердцевину. Пузырьки образуются путем отпочковывания от агранулярной
26
эндоплазматической сети, а содержащиеся в пероксисомах ферменты — в гЭПС. Пероксисомы выявлены в клетках печени, почек и в других органах.
Мембрана пероксисом обладает высокой проницаемостью для ионов и веществ с низкой молекулярной массой. Матрикс содержит ряд ферментов, наиболее значимыми из которых являются пероксидаза, каталаза, оксидаза для окисления аминокислот и уратоксидаза. Сердцевина пероксисом — это область конденсации ферментов, которая состоит из фибрилл и трубочек и имеет кристаллоидную структуру.
Функции пероксисом. Пероксисомы осуществляют реакции детоксикации в клетке: распад перекиси водорода, которая образуется в клетках в результате окисления аминокислот, углеводов и других веществ и является токсичной; окисление этилового спирта; катализируют расщепление жирных кислот и др. Генетические нарушения ферментной активности пероксисом ведут к тяжелым повреждениям нервной системы (пероксисомные болезни).
10. Энергетическийаппаратклетки: митохондрии. Структурнаяорганизация, типымитохондрий. Митохондриальный матрикс. Жизненный цикл митохондрий
Митохондрии — двухмембранные клеточные органеллы, обеспечивающие клетку энергией, запасаемой в виде фосфатных связей АТФ, участвующие в синтезе стероидов, окислении жирных и нуклеиновых кислот. Основная функция — окисление органических веществ и использование освобождающейся при этом энергии для нужд клетки.
Форма митохондрий различна, их размеры колеблются: в ширину от 0,2 до 2 мкм, в длину — от 2 до 10 мкм. Число митохондрий зависит от активности клетки. В клетке они располагаются, как правило, в зонах максимального функционирования И потребления энергии в области синтетического и двигательного аппаратов клетки, вблизи ионных каналов.
Строение. Митохондрии образованы двумя мембранами — наружнойивнутренней. Мембраныразделенымежмембраннымпространством, содержащим ферменты. (Рис. 5).
Наружная мембрана организована по принципу цитолеммы. Она обладает высокой проницаемостью и содержит молекулы транспортных белков, на ее поверхности содержатся рецепторы.
Внутренняямембранаобладаетнизкойпроницаемостьюдлямелкихионов, содержиттранспортныебелки, ферментыдыхательной
27
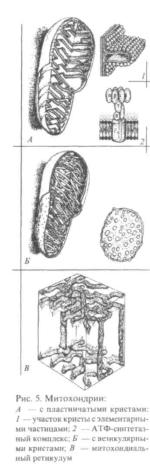
цепи |
и |
АТФ-синтетазные |
комп- |
лексы. Внутренняямембранаоб- |
|
||
разует складки, идущие перпендикулярно длинной оси органеллы — кристы, количество которых в одной митохондрии составляет несколько десятков.
Кристы внутренней мембраны митохондрий у большинства клеток имеют пластинчатую форму и лишь у некоторых клеток содержатся везикулярные или трубчатые кристы (в эндокринных клетках, продуцирующих стероидные гормоны).
На кристах расположены элементарные грибовидные частицы, обеспечивающие сопряжение процессов окисления и фосфорилирования, осуществление синтеза АТФ (АТФ-синтетазные комплексы). При распаде фосфатных связей АТФ освобождается большое количество энергии, используемое для обеспечения биосинтетических процессов в клетке.
Митохондриальный матрикс,
заполняющий внутреннюю полость митохондрий, является веществом средней электронной плотности и содержит митохон-дриальные (1) ферменты, (2) гра-
нулы, (3) рибосомы и (4) ДНК.
Ферменты участвуют в обеспечении цикла Кребса, окислении жирных кислот, синтезе белков. Гранулы представлены округлыми частицами высокой электронной плотности, содержат ионы Са2+ и Мg2+, необходимые для поддержания активности ферментов. Рибосомы присутствуют в матриксе в виде мелких плотныхчастиц.
28
Особенностью митохондрий является наличие в них собственного генома, представленного митохондриальной ДНК (мтДНК), которая включает 37 генов. На долю мтДНК приходится 1% от всей ДНК, содержащейся в клетке. Митохондриальный геном осуществляет синтез 5% белков, в основном электронно-транспортной системы и ферментов синтеза ЛТФ. МтДНК наследуется только по материнской линии. Поскольку в матриксе митохондрий содержится много перекиси водорода и активных радикалов кислорода, часто встречаются мутации мтДНК, являющиеся основой ряда заболеваний (митохондриальные болезни).
Жизненный цикл митохондрий составляет примерно 10
суток. По окончании этого периода времени они подвергаются разрушению аутофагией.
Новообразование митохондрий осуществляется перешнуровкой зрелых органелл; их делением.
Митохондрии функционируют во взаимосвязи с эндоплазматической сетью, обеспечивают синтез белков в рибосомах и являются силовыми станциями клетки. Они очень чувствительны к изменениям метаболизма клетки — трофики, рН среды, осмотического давления. Митохондрии первыми среди клеточных органелл реагируют на воздействия повреждающих факторов.
11.Включения, их классификация, химическая
иморфофункциональная характеристика
Клеточные включения — это непостоянные компоненты клетки, которые синтезируются или запасаются в виде гранул в цитоплазме клеток различных тканей в зависимости от специфики обмена веществ.
Классификация включений. Включения могут быть эндогенными и экзогенными. Их классифицируют по химическому составу и функциональному назначению.
По химическому составу различают включения: липидные, белковые, углеводные, сложного химического состава.
По функциональному назначению выделяют: трофические,
секреторные, экскреторные, пигментные.
Примеры конкретных включений в разных клетках: для яйцеклетки характерны включения белка; для гепатоцитов — гликогена; жировые включения накапливаются в липоцитах соеди-тельной ткани; пигментные (гранулы меланина) — в клетках эпидермиса, оболочках глаза.
Морфофункциональная характеристика. Включения,
образован-преимущественно жидкостным компонентом, представлены в
29
цитоплазме в виде капель, называемых вакуолями. Если включения содержат плотный компонент, их именуют гранулами.
Трофические включения (жировые, углеводные, белковые) необходимы клетке для обменных процессов. Их запас может определять уровень функционирования клетки.
Секреторные — содержат вещества, синтезированные в самой клетке и обладающие, в ряде случаев, биологической активностью (гормоны, проферменты).
Пигментные включения могут быть экзогенными по природе (каротин, красители, пылевые частицы), но чаще — эндогенными (гемоглобин эритроцитов, меланин, липофусцин — пигмент старения). Их присутствие в клетке может придавать ткани или органу специфический цвет, как в норме (цвет кожи, волос), так и при патологии (желтушная окраска кожи при наличии билирубина). В последнем случае это может иметь диагностическое значение.
Экскреторные включения — это продукты обмена, подлежащие удалению из клетки.
Не являясь постоянной составной частью цитоплазмы, включения отражают уровень и особенности обмена веществ в клетке, т. е. имеют определенное значение для ее жизнедеятельности.
12.Ядро: основные компоненты
иих структурно-функциональная характеристика. Ядерно-цитоплазматические отношения как показатель функционального состояния клеток
Ядро — важнейший и обязательный структурный компонент эукариотической клетки. Оно обеспечивает:
—хранение и передачу генетической информации;
—создаёт аппарат белкового синтеза (на молекулах ДНК идёт транскрипция информационных, транспортных и Рибосомных РНК);
—участвует в регуляции и координации деятельности всех цитоплазматических структур.
Клетки без ядер не образуют белок и имеют ограниченную активность. Форма и размеры ядер зависят от формы и функции клеток.
Строение. Ядро состоит из хроматина, ядрышка, кариоплазмы и ядерной оболочки. (Рис. 6).
Хроматин. В состав хроматина входит ДНК в комплексе с белком. В интерфазных ядрах он представляет собой хромосомы, которые в это время разрыхляются, деконденсируются. Зоны
30