

Gistologia_v_voprosakh_i_otvetakh_Sluka
.pdf(по краям мешает костная манжетка). Кровеносные сосуды с мезенхимой, остеобластами и остеокластами врастают через отверстия костной манжетки в диафизарный хрящ. Остеокласты его разрушают, а остеобласты с остеоцитами формируют костную ткань (эндохондральные окостенение).
В последующем две зоны окостенения — перихондральная и энхондральная сливаются вместе. Внутри диафиза формируется костномозговая полость, которая заселяется мислоидной тканью. Вслед за диафизом центры окостенения появляются в эпифизах, что сопровождается врастанием в эпифиз сосудов. Постэмбриональное развитие кости осуществляется при регенерации и эктопическом остеогенезе.
Гл а в а 5. Мышечные ткани
44.Мышечные ткани. Общая морфофункциональная характеристика. Классификация. Источники развития. Регенерация мышечных тканей
Мышечные ткани выполняют двигательные функции и обеспечивают перемещение организма в целом и его частей в пространстве, так как элементы этих тканей способны сокращаться изменять свою форму под влиянием пусковых (нервных) импульсов. Сокращение — результат взаимодействия молекул сократительных белков (актина и миозина), которое происходит с участием других белков и ионов кальция
(акто-миозиновый механохимический преобразователь). Для сокращения требуется энергия, поэтому структурные элементы мышечных тканей богаты митохондриями. В их цитоплазме мно-го гликогена и миоглобина.
В структурно-функциональной организации
мышечных элементов клеток (миоцитов) и мышечных волокон (миосимпластов) удобно выделить три аппарата:
1)опорный — двухслойная сарколемма с сетью эластических волокон в базальной мембране;
2)трофический — саркоплазматическая сеть, кавеолы или пиноцитозные везикулы, митохондрии, включения гликогена; 3) сократительный — миофибриллы.
Классификация. Согласно морфофункциональной классифика-ции мышечные ткани подразделяются на гладкие (неисчерченные) и перечнополосатые (исчерченные). В цитоплазме клеток гладкиз мышечных тканей актиновые и миозиновые миофиламенты
101
(протофибриллы) образуют миофибриллы, не имеющие поперечной исчерченности. Мышечные элементы поперечнополосатых тканей постоянно содержат в цитоплазме и актиновые, и миозиновые протофибриллы, взаимное расположение которых в составе миофибрилл создает поперечную исчерченность.
Источники развития. В зависимости от источника развития мышечные ткани делятся на 1) мезенхимные (гладкие мышцы внутренних органов и сосудов), 2) эпидермальные (миоэпите-лиальные клетки), 3) нсйральные (мышцы, суживающие и расширяющие зрачок), 4) целомические (сердечная мышца) и 5) соматические (миотомные, скелетные). Первые три типа относятся к гладким мышечным тканям, четвертый и пятый — к поперечнополосатым.
Регенерация. Для гладкой мышечной ткани характерна выраженная физиологическая и репаративная регенерация, которая проявляется как в форме компенсаторной гипертрофии, так и клеточной регенерации. Камбием для нее служат стволовые и адвентициальные клетки рыхлой соединительной ткани. Глад-комышечные клетки (прежде всего малодифференцированные) способны делиться митозом, за счет чего происходит постоянное обновление клеточного состава гладких мышц и постоянная их перестройка в ряде органов (матка, сосуды и др.). Подобно фибробластам, гладкомышечные клетки способны образовывать межклеточное вещество.
Восстановление волокон скелетной мышечной ткани идет за счет реактивных изменений сохранившейся части волокна и размножения камбиальных клеток — миосателлитоцитов. На первых этапах в поврежденных мышечных волокнах происходит деструкция миофибриллярного аппарата. В сохранившихся участках активируются процессы синтеза РНК и белков. В области повреждения за счет активизации и дифферен-цировки клеток-сателлитов образуются многочисленные миобласты. В дальнейшем миобласты сливаются в миосимп-ласты, последние преобразуются в мышечные трубочки и далее — в зрелые мышечные волокна.
Для исхода регенерации существенное значение имеют оптимальные межтканевые взаимодействия мышечной ткани с нервной и соединительной, механическое напряжение и кровоснабжение.
Сердечная мышечная ткань не сохраняет ни стволовых клеток, ни клеток-предшественников, а кардиомиоциты находятся в фазе С0, в связи с чем регенерация кардиомиоцитов невозможна и, если кардиомиоциты гибнут (инфаркт), то они не восстанавливаются.
102
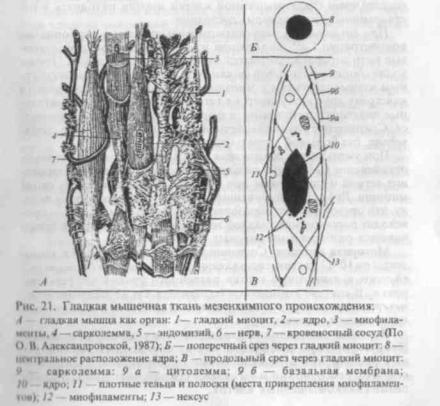
45. Гладкие мышечные ткани по источникам развития делятся на мезенхимные, эпидермальные и нейральные. Все они имеют клеточное строение. Основной гистологический элемент ткани — глад-комышечная клетка, способная к гипертрофии и регенерации, а также к синтезу межклеточного вещества. Гладкие мышцы образуют мышечные оболочки полых и трубчатых органов, кровеносных сосудов, регулируя
величину их просвета и моторику. Регуляцию деятельности гладкой мышечной ткани осуществляют вегетативная нервная системаигуморальныефакторы(рис.21)
103
Гладкие миоциты имеют веретенообразную или звездчатую форму, длиной от 20 мкм до 1 мм. Их базалъная мембрана содержит протеогликаны, коллаген типа III и V. Компоненты базаль-ной мембраны и эластин межклеточного вещества синтезируются как самими миоцитами, так и фибробластами соединительной ткани. Ядро расположено в центре клеток, форма его эллипсоидная. В саркоплазме у полюсов ядра находятся многочисленные митохондрии, развитый комплекс Гольджи, саркоплазма-тический ретикулум и свободные рибосомы. Миофиламенты ориентированы преимущественно вдоль длинной оси клетки, они обеспечивают ее сокращение.
Механизм сокращения. В цитоплазме миоцитов постоянно присутствуют только актиновые миофиламенты; миозиновые выявляются в виде нитей только в сокращенном состоянии. При расслаблении гладкомышечной клетки миозин находится в растворенном (разобранном) состоянии.
При сокращении, запускаемом повышением в саркоплазме концентрации Са2+, выходящем из депо, актиновые и миозиновые нити перераспределяются относительно друг друга. Происходит сборка миозиновой филаменты. Благодаря межмолекулярным взаимодействиям с миозином актиновые нити смещаются навстречу друг другу, энергия тяги передается на прикрепительные пластинки цитолеммы, и форма клетки при этом изменяется. Сокращение гладкомышечной ткани непроизвольное, тоническое, без большой затраты энергии.
При уменьшении содержания Са2* в саркоплазме (постоянное откачивание Са2+ в депо кальция) происходит дефосфорилирова-ние легкой цепи миозина при помощи фосфатазы легких цепей миозина. Дефосфорилированный миозин теряет сродство к актину, что предотвращает образование поперечных мостиков — происходит расслабление гладкой мышечной клетки, которое заканчивается разборкой миозиновых филаментов.
Моторная единица. Соседние миоциты объединены в комплексы по 10-12 клеток, составляющие эффекторную (моторную) единицу, включающую клетки различного уровня дифференци-ровки. В составе комплексов содержатся также миоциты-пейс-мекеры, сами генерирующие потенциал действия и передающие его соседним клеткам.
Нервные окончания подходят ко всему такому комплексу. Нейромедиаторы диффундируют из варикозных терминальных расширений в межклеточные пространства, взаимодействуют с рецепторами и вызывают сокращение, либо расслабление гладкомышечных клеток.
104
Гладкомышечные клетки функционируют не как изолированные единицы, а в виде клеточных комплексов. Это достигается с помощью щелевидных контактов (нексусов), обеспечивающих взаимодействие миоцитов. Возбуждение клеток, не имеющих иннервации, происходит либо за счет медленной диффузии нейромедиатора, либо посредством щелевых контактов между клетками.
Гладкая мышца как орган. Вокруг мышечных клеток ретикулярные, эластические и тонкие коллагеновые волокна образуют трехмерную сеть — эндомизий. В составе органов гладкие миоциты объединяются в пучки, между которыми располагаются тонкие прослойки рыхлой волокнистой соединительной ткани (перимизий), а совокупность пучков образуют мышцу, которая окружена более толстыми прослойками волокнистой соединительной ткани (эпимизий).
Вних проходят кровеносные сосуды и нервные волокна. Практически вся мышечная ткань внутренних органов
является гладкой мышечной тканью мезенхимного происхождения (за исключением языка, гортани, верхних отделов пищевода).
Мионейральная ткань. Миоциты мышечной ткани нейраль-ного происхождения (миопигментоциты, мионейральные) развиваются из клеток нейрального зачатка в составе стенки глазного бокала. Они входят в состав двух мышц радужки глаза — расширяющей и суживающей зрачок. Это одноядерные верете-новидные клетки, содержащие миофибриллы (тонкие — 7 нм и толстые — 15 нм), большое число митохондрий и пигментные гранулы. Между клетками имеются щелевые контакты (нексусы), плотные и десмосомоподобные соединения. Мышцы получают вегетативную иннервацию — двигательные нервные окончания подходят к каждой мионейральной клетке. Мышца, расширяющая зрачок, получает симпатическую иннервацию (пещеристое сплетение); мышца суживающая зрачок — парасимпатическую, от постганглионарных нейронов ресничного ганглия.
Немышечные сокращающиеся клетки. Элементами мышечной ткани эпидермального происхождения являются
миоэпителиальные клетки, развивающиеся из кожной эктодермы. Они имеют звездчатую форму и охватывают отростками концевые отделы и мелкие выводные протоки желез (слюнных, молочных, слезных, потовых). Стабильные актиновые миофиламенты, прикрепленные к плотным тельцам, и нестабильные миозиновые миофиламенты, формирующиеся в процессе сокращения, образуют сократительный аппарат миоэпителиальных клеток. Сокращаясь, эти клетки способствуют выведению секрета желез. Холинергические нервные волокна стимулируют сокращение
105
миоэпителиальных клеток слюнных и слезных желез, окситоцин — лактирующих молочных желез.
Миофибробласты, способные синтезировать сократительные белки (актин и миозин) при заживлении раны, способствуют сближению раневых поверхностей.
46.Поперечнополосатая скелетная мышечная ткань. Строение, иннервация. Структурныеосновы сокращениямышечного волокна. Типы мышечных волокон. Строение мышцы как органа
Скелетная поперечнополосатая мышечная ткань
составляет около 40% массы тела человека и обеспечивает осознанные и осознаваемые произвольные движения тела и его частей. Основными структурными элементами являются скелетные мышечные волокна (миосимпласты), выполняющие функцию сокращения, и миосателлиты (камбиальный резерв).
Источником развития являются миотомы (части сомитов мезодермы). Миогистогенез — процесс развития мышечных волокон включает ряд стадий:
1)миобластическая (миграция в места закладок скелетных мышц одноядерных верстеновидных митотически делящихся миобластов);
2)миосимпластическая (миобласты располагаются в виде цепочки вдоль ориентированных между скелетными закладками колла-геновых волокон и сливаются друг с другом в миосимпласты);
3)миотубулярная (в миосимпластах формируются поперечно-полосатые миофибриллы, которые вначале располагаются по периферии, а ядра в центре);
4)мышечных волокон (миофибриллы постепенно заполняют весь объем саркоплазмы, смещая ядра на периферию, под сарколемму).
Строение. Построена из мышечных волокон. К волокну прилежит клетка — миосателлитоцит, имеющая гиперхромное ядро и общую с волокном оболочку (сарколемму), которая образована базальной мембраной и плазмолеммой симпласта. Миосателлито-циты прилежат к поверхности симпласта так, что их плазмалеммы соприкасаются. С одним симпластом связано много сателлитоци-тов. Каждый миосателлитоцит — одноядерная клетка — является камбиальным элементом скелетной мышечной ткани. (Рис. 22)
Мышечные волокна — структуры цилиндрической формы длиной от нескольких мм до 10 см, имеют множество ядер, расположенных под сарколеммой. В цитоплазме (саркоплазме) много саркосом (митохондрий) и канальцев эндоплазматической сети
106
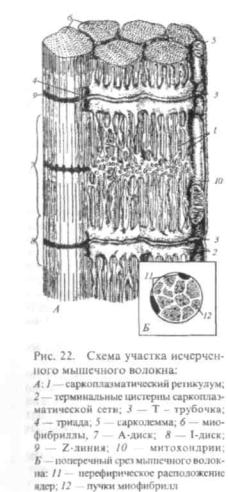
(гладкая сеть образует систему Т- и L-трубочек, депонирующих ионы кальция).
Миофибриллы располагаются параллельно оси волокна от одного его конца до другого. Они состоят из чередующихся участков с разными физико-хи- мическими и оптическими свойствами. Разный коэффициент преломления света в этих участках обусловливает деление миофибрилл на темные и светлые сегменты (диски А
— темные, анизотропные; диски И — светлые, изотропные). За эту исчерченность миофибрилл мышечные волокна получили
название поперечнополосатых. Через середину дисков И от оболочки волокна проходят мембраны Т (телофрагмы).
Участок миофибриллы между двумя телофрагмами называется саркомером.
В состав миофибрилл входят миофиламенты, состоящие из белков: тонкие— из актина, тропонина, тропомиози-на; толстые — из миозина.
Иннервация. Нервы,
вступающие в мышцу, содержат как эфферентные (двигательные, аксоны а- и g -мотонейронов переднего рога спинного мозга и двигательных ядер
черепных нервов), так и афферентные (чувствительные, денд-риты нейронов сиинальных ганглиев и чувствительных узлов черепных нервов) волокна.
Соответственно характеру иннервации выделяются два типа мышечных волокон: экстрафузальные и интрафу зальные.
Каждое экстрафузальное мышечное волокно иннервирустся самостоятельно терминальными ветвлениями
аксонов
107
алъфа-мотонейронов с образованием нервно-мышечных синапсов. Один мотонейрон и иннервируемая им группа экстрафу-зальных мышечных волокон образует морфофункциональную нейромоторную единицу скелетной мышцы — мион.
Интрафузальные мышечные волокна образуют нервномышечные синапсы с эфферентными волокнами гаммамотонейронов и вместе с чувствительными нервными окончаниями формируют нервно-мышечные веретена — рецепторы скелетной мышцы.
Вегетативная иннервация мышечных волокон не обнаружена, но гладкие миоциты кровеносных сосудов мышц имеют симпатическую адренергическую иннервацию.
Структурные основы сокращения под влиянием потенциала действия, который распространяется по цитолемме и Т-трубочкам, ионы кальция выходят в саркоплазму, соединяются с белком тро-понином, который при этом изменяет конформацию. Так как тро-понин в составе тонких нитей связан с тропомиозином, конформация тропомиозина также изменяется, его молекула отодвигается и открывает активные центры актина, благодаря которым актино-вые нити соединяются с миозиновыми, образуя актомиозиновый комплекс. Тонкие миофиламенты скользят между толстыми, сокращение происходит одновременно во всех саркомерах миофибриллы. Максимально длина саркомера может уменьшиться на 1 /3 часть от первоначальной в состоянии расслабления. При расслаблении ионы кальция возвращаются обратно в Т- цистсрны.
Типы мышечных волокон различаются на основе характера сокращения, скорости сокращения и типа окислительного обмена.
По характеру сокращения экстрафузальные мышечные волокна разделяются на фазные (отвечающие на нервную стимуляцию быстрым сокращением) и тонические (имеющие более низкий потенциал покоя и сокращающиеся после повторных нервных стимулов). Тонические мышцы (ушные и наружные глазные мышцы) имеют полинейронную иннервацию.
По скорости сокращения, определяемой активностью АТФ-фазы миозина, выделяются быстрые и медленные мышечные волокна.
По типу окислительного обмена :
1)окислительные или красные мышечные волокна небольшого диаметра, содержащие много миоглобина, окруженные массой капилляров и имеющие в митохондриях высокий уровень активности окислительных ферментов (быстрые неутомляемые и медленные неутомляемые);
2)гликолитические или белые, имеющие больший диаметр, со значительным количеством гликогена, немногочисленными митохондриями, с низкой активностью окислительных и
108
высокой — гликолитических ферментов (быстро сокращающиеся и быстроутомляемые). Различие цвета красных и белых волокон связано с разным содержанием в них белка миоглобина, способного связывать кислород и при необходимости медленно отдавать его мышечной ткани;
3) промежуточные (окислительно-гликолитические) мышечные волокна - с умеренной активностью окислительных ферментов, низкой активностью гликогена, медленно утомляемые.
Строение мышцы как органа. В зависимости от преобладания определенного типа мышечных волокон скелетные мышцы делятся на красные (медленные), быстрые (белые), либо смешанные. В мышце как органе между мышечными волокнами находятся тонкие прослойки рыхлой волокнистой соединительной ткани — эндомизий. Мышечные волокна собираются в пучки, между которыми лежат более толстые соединительнотканные прослойки — перимизий. Снаружи мышца покрыта эпимизием (фасцией), образованным волокнистой соединительной тканью. Концы мышцы переходят в сухожилия.
47. Поперечно-полосатая сердечная мышечная ткань. Структурно-функциональная характеристика. Источники развития и регенерация
Сердечная поперечно-полосатая мышечная ткань
развивается из симметричных участков висцерального листка спланхното-ма (целомической выстилки) в шейной части тела зародыша, называемых миоэпикардиальной пластинкой. В ходе гистогенеза образуется несколько видов кардиомиоцитов: сократительные, проводящие и секреторные. Сократительные клетки имеют удлиненную форму, их концы соединяются друг с другом так, что цепочки кардиомиоцитов составляют функциональные волокна, а области контакта образуют вставочные диски. (Рис.23).
Во вставочных дисках выделяют межфибриллярный отдел
(поперечный участок, где образуются десмосомы с прикрепленными к ним тонкими миофиламентами миофибрилл и интердигитации) и меж-саркоплазматическш (боковые, продольные части, где преобладают щелевые контакты — нексусы, обеспечивающие сокращение множества кардиомиоцитов в составе функционального синцития).
Кардиомиоциты могут ветвиться и образуют пространственную сеть. Их наружные поверхности покрыты базальной мембраной. Ядро (или ядра, если их два) овальное, лежит в центральной части клетки. Агранулярная эндоплазматическая сеть хорошо развита. Она формирует цистерны, прилежащие к Т-системам,
109
