

- •Contents
- •Preface to the first edition
- •Flagella
- •Cell walls and mucilages
- •Plastids
- •Mitochondria and peroxisomes
- •Division of chloroplasts and mitochondria
- •Storage products
- •Contractile vacuoles
- •Nutrition
- •Gene sequencing and algal systematics
- •Classification
- •Algae and the fossil record
- •REFERENCES
- •CYANOPHYCEAE
- •Morphology
- •Cell wall and gliding
- •Pili and twitching
- •Sheaths
- •Protoplasmic structure
- •Gas vacuoles
- •Pigments and photosynthesis
- •Akinetes
- •Heterocysts
- •Nitrogen fixation
- •Asexual reproduction
- •Growth and metabolism
- •Lack of feedback control of enzyme biosynthesis
- •Symbiosis
- •Extracellular associations
- •Ecology of cyanobacteria
- •Freshwater environment
- •Terrestrial environment
- •Adaption to silting and salinity
- •Cyanotoxins
- •Cyanobacteria and the quality of drinking water
- •Utilization of cyanobacteria as food
- •Cyanophages
- •Secretion of antibiotics and siderophores
- •Calcium carbonate deposition and fossil record
- •Chroococcales
- •Classification
- •Oscillatoriales
- •Nostocales
- •REFERENCES
- •REFERENCES
- •REFERENCES
- •RHODOPHYCEAE
- •Cell structure
- •Cell walls
- •Chloroplasts and storage products
- •Pit connections
- •Calcification
- •Secretory cells
- •Iridescence
- •Epiphytes and parasites
- •Defense mechanisms of the red algae
- •Commercial utilization of red algal mucilages
- •Reproductive structures
- •Carpogonium
- •Spermatium
- •Fertilization
- •Meiosporangia and meiospores
- •Asexual spores
- •Spore motility
- •Classification
- •Cyanidiales
- •Porphyridiales
- •Bangiales
- •Acrochaetiales
- •Batrachospermales
- •Nemaliales
- •Corallinales
- •Gelidiales
- •Gracilariales
- •Ceramiales
- •REFERENCES
- •Cell structure
- •Phototaxis and eyespots
- •Asexual reproduction
- •Sexual reproduction
- •Classification
- •Position of flagella in cells
- •Flagellar roots
- •Multilayered structure
- •Occurrence of scales or a wall on the motile cells
- •Cell division
- •Superoxide dismutase
- •Prasinophyceae
- •Charophyceae
- •Classification
- •Klebsormidiales
- •Zygnematales
- •Coleochaetales
- •Charales
- •Ulvophyceae
- •Classification
- •Ulotrichales
- •Ulvales
- •Cladophorales
- •Dasycladales
- •Caulerpales
- •Siphonocladales
- •Chlorophyceae
- •Classification
- •Volvocales
- •Tetrasporales
- •Prasiolales
- •Chlorellales
- •Trebouxiales
- •Sphaeropleales
- •Chlorosarcinales
- •Chaetophorales
- •Oedogoniales
- •REFERENCES
- •REFERENCES
- •EUGLENOPHYCEAE
- •Nucleus and nuclear division
- •Eyespot, paraflagellar swelling, and phototaxis
- •Muciferous bodies and extracellular structures
- •Chloroplasts and storage products
- •Nutrition
- •Classification
- •Heteronematales
- •Eutreptiales
- •Euglenales
- •REFERENCES
- •DINOPHYCEAE
- •Cell structure
- •Theca
- •Scales
- •Flagella
- •Pusule
- •Chloroplasts and pigments
- •Phototaxis and eyespots
- •Nucleus
- •Projectiles
- •Accumulation body
- •Resting spores or cysts or hypnospores and fossil Dinophyceae
- •Toxins
- •Dinoflagellates and oil and coal deposits
- •Bioluminescence
- •Rhythms
- •Heterotrophic dinoflagellates
- •Direct engulfment of prey
- •Peduncle feeding
- •Symbiotic dinoflagellates
- •Classification
- •Prorocentrales
- •Dinophysiales
- •Peridiniales
- •Gymnodiniales
- •REFERENCES
- •REFERENCES
- •Chlorarachniophyta
- •REFERENCES
- •CRYPTOPHYCEAE
- •Cell structure
- •Ecology
- •Symbiotic associations
- •Classification
- •Goniomonadales
- •Cryptomonadales
- •Chroomonadales
- •REFERENCES
- •CHRYSOPHYCEAE
- •Cell structure
- •Flagella and eyespot
- •Internal organelles
- •Extracellular deposits
- •Statospores
- •Nutrition
- •Ecology
- •Classification
- •Chromulinales
- •Parmales
- •Chrysomeridales
- •REFERENCES
- •SYNUROPHYCEAE
- •Classification
- •REFERENCES
- •EUSTIGMATOPHYCEAE
- •REFERENCES
- •PINGUIOPHYCEAE
- •REFERENCES
- •DICTYOCHOPHYCEAE
- •Classification
- •Rhizochromulinales
- •Pedinellales
- •Dictyocales
- •REFERENCES
- •PELAGOPHYCEAE
- •REFERENCES
- •BOLIDOPHYCEAE
- •REFERENCE
- •BACILLARIOPHYCEAE
- •Cell structure
- •Cell wall
- •Cell division and the formation of the new wall
- •Extracellular mucilage, biolfouling, and gliding
- •Motility
- •Plastids and storage products
- •Resting spores and resting cells
- •Auxospores
- •Rhythmic phenomena
- •Physiology
- •Chemical defense against predation
- •Ecology
- •Marine environment
- •Freshwater environment
- •Fossil diatoms
- •Classification
- •Biddulphiales
- •Bacillariales
- •REFERENCES
- •RAPHIDOPHYCEAE
- •REFERENCES
- •XANTHOPHYCEAE
- •Cell structure
- •Cell wall
- •Chloroplasts and food reserves
- •Asexual reproduction
- •Sexual reproduction
- •Mischococcales
- •Tribonematales
- •Botrydiales
- •Vaucheriales
- •REFERENCES
- •PHAEOTHAMNIOPHYCEAE
- •REFERENCES
- •PHAEOPHYCEAE
- •Cell structure
- •Cell walls
- •Flagella and eyespot
- •Chloroplasts and photosynthesis
- •Phlorotannins and physodes
- •Life history
- •Classification
- •Dictyotales
- •Sphacelariales
- •Cutleriales
- •Desmarestiales
- •Ectocarpales
- •Laminariales
- •Fucales
- •REFERENCES
- •PRYMNESIOPHYCEAE
- •Cell structure
- •Flagella
- •Haptonema
- •Chloroplasts
- •Other cytoplasmic structures
- •Scales and coccoliths
- •Toxins
- •Classification
- •Prymnesiales
- •Pavlovales
- •REFERENCES
- •Toxic algae
- •Toxic algae and the end-Permian extinction
- •Cooling of the Earth, cloud condensation nuclei, and DMSP
- •Chemical defense mechanisms of algae
- •The Antarctic and Southern Ocean
- •The grand experiment
- •Antarctic lakes as a model for life on the planet Mars or Jupiter’s moon Europa
- •Ultraviolet radiation, the ozone hole, and sunscreens produced by algae
- •Hydrogen fuel cells and hydrogen gas production by algae
- •REFERENCES
- •Glossary
- •Index
HETEROKONTOPHYTA, BACILLARIOPHYCEAE |
395 |
|
|
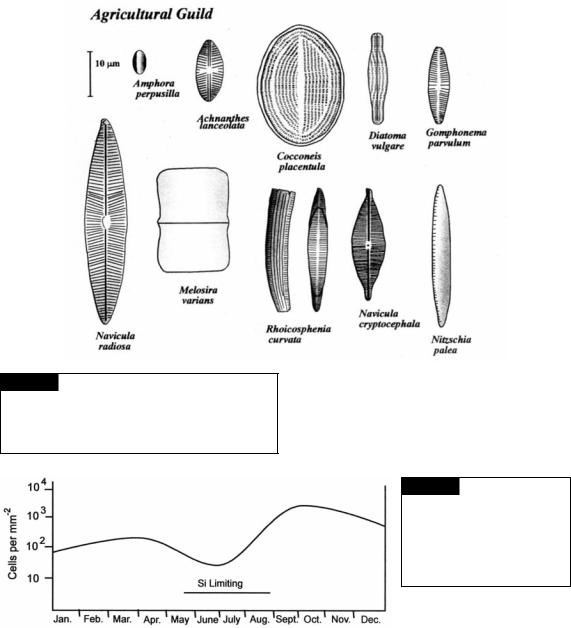
Fig. 17.35 Diatoms that are characteristic of eutrophic freshwaters. The diatoms make up the “agricultural guild”, named for waters that are polluted by runoff from agricultural land that have been fertilized with nitrogen, phosphorus, and potash.
Fig. 17.36 Seasonal fluctuations of epiphytic Tabellaria flocculosa on
Phragmites communis and
Schoenoplectus lacustris stems. The straight line represents the period when dissolved silica is less then 0.5 mg liter 1. (After Knudson, 1957.)
In freshwater habitats, diatoms often comprise the dominant algal flora in thermal waters between 30 and 40 °C. Fairchild and Sheriden (1974) showed that Achnanthes exigua (Fig. 17.33(f )) isolated from a hot spring showed optimum photosynthesis at 42 °C, with maximum and minimum temperatures for growth at 44 and 10 °C, respectively – characteristics of a thermophilic organism. This particular diatom is usually associated with an
algal mat covered by a shallow layer of water in these hot springs.
Fossil diatoms
The fossil record of diatoms begins in the Jurassic Period (185 million years ago) with diatoms becoming more common in the mid-Cretaceous
396 CHLOROPLAST E.R.: EVOLUTION OF TWO MEMBRANES
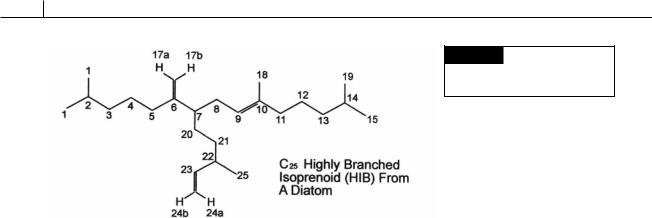
Fig. 17.37 Chemical structure of
a highly branched alkene
characteristic of diatoms.
(Medlin et al., 1997). The fossil record indicates that centric diatoms evolved before pinnate diatoms (Round et al., 1990). Another way of dating diatoms is by molecular fossils. Some diatom genera (Rhizosolenia, Fig. 17.33(c); Haslea, Navicula, Figs. 17.2(d), 17.16, 17.26(a), 17.33(b), 17.35; and Pleurosigma) contain unique isoprenoids called highly branched isoprenoid (HBI) alkenes that have a T-branch formed by attachment of a C10 isoprenoid unit to a C15 isoprenoid unit (Fig. 17.37) (Damste et al., 2004). The occurrence of these alkenes in sediments beginning 90 million years ago parallels the widespread explosion of diatoms in the marine environment at this time.
After death of diatom cells, the frustule usually dissolves after bacteria degrade the organic casing around the frustule (Bidle and Azam, 1999). However, under certain circumstances, the frustules remain intact and accumulate at the bottom of the water (Fig. 17.38). Where conditions are exceptionally favorable and long continued, such accumulations may reach considerable thickness. Deposits of fossil diatoms, known as diatomaceous earth or kieselguhr, are found in various parts of the world. None of these deposits originated earlier than the Cretaceous; some diatomaceous earths originated in freshwaters, others in the ocean. However, known deposits of marine species are found inland and above the ocean as a result of geological changes. The best-known and most extensive deposits of marine species are those at Lompoc, California, where the beds are several kilometers in extent and 200 m in thickness (Fig. 17.39). Most of the commercially available diatomaceous earth comes from California, where the deposits are worked as open quar-
ries. In quarrying, the overburden of soil is removed, and the diatomaceous earth is mined. Diatomaceous earth is also obtained from lakes in Florida by dredging with a suction pump and carrying the material through sluiceways to settling tanks. The material from some deposits can be used directly; that from other deposits must be incinerated to remove organic substances.
The industrial uses of diatomaceous earth are varied. One of the first uses was as a mild abrasive in toothpaste and metal polishes. Diatomaceous earth was also used as an absorbent for liquid nitroglycerin to make dynamite that could be transported with comparative safety. The inert medium used in the present-day manufacture of dynamite is wood meal. Probably the most extensive industrial use of diatomaceous earth is in the filtration of liquids, especially those of sugar refineries. Another major use is in the insulation of boilers, blast furnaces, and other places where a high temperature is maintained.
Analysis of sediments containing remains of diatoms can provide information on past environmental conditions of lakes (Reavic et al., 1998; Clavero et al., 2000). This technique has been applied to late glacial and postglacial diatom deposits in the English Lake District (Round, 1957, 1961). During the end of the last glacial period, there were few diatoms present, with the sediments being mostly of inorganic origin, indicating that the lakes initially were very oligotrophic and lacked planktonic diatoms. This period was followed by diatoms characteristic of melt waters under semi-arctic conditions (Melosira arenaria, Fig. 17.40(a). Cyclotella antique, Fig. 17.40(b); Gyrosigma, Fig. 17.40(d); Cymatopleura, Fig. 17.40(e);
HETEROKONTOPHYTA, BACILLARIOPHYCEAE |
397 |
|
|
Fig. 17.38 Scanning electron micrographs of fossil diatoms compared with frustules of living diatoms from the same genera. (a)
Coscinodiscus radiatus from
Cretaceous sediments. (b)
Coscinodiscus radiatus from recent plankton. (c) Stephanopyxis turris from Cretaceous sediments.
(d) Stephanopyxis broschi from recent plankton. (From Medlin et al., 1993.)
Fig. 17.39 Diatomaceous earth mine at Lompoc,
California, with deposits of diatomaceous earth in the
background. (Courtesy of Johns-Manville Corporation.)