Chapter 9
Cryptophyta
CRYPTOPHYCEAE |
and Lee, 1987). The number and shape of these |
|
plates are used to characterize genera taking into |
||
|
consideration that the haploid and diploid phases |
|
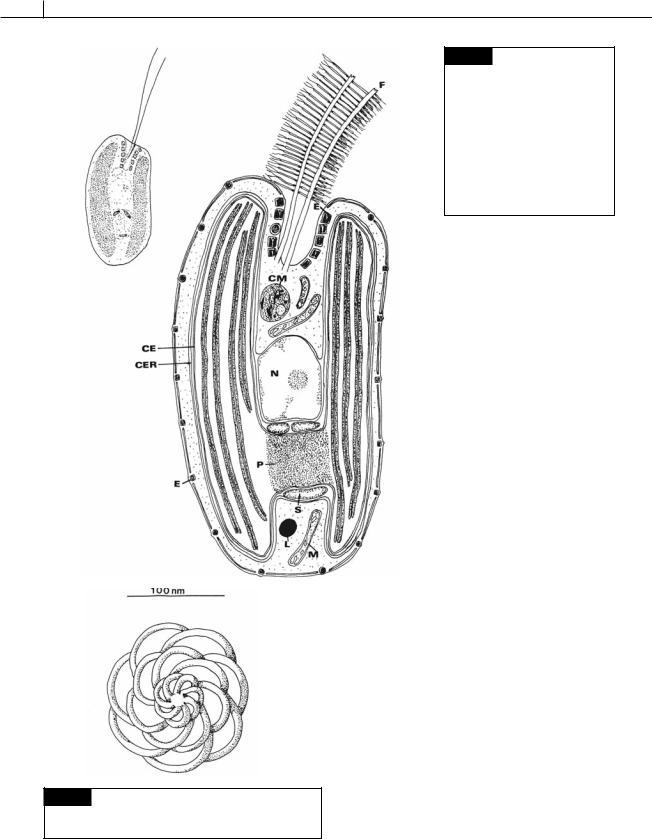
This group is composed primarily of flagellates |
of a single genus can have different plates (Hoef- |
|
that occur in both marine and freshwater envir- |
Emden and Melkonian, 2003). New periplast plates |
|
onments. The cells contain chlorophylls a and c2 |
are added in an area adjacent to the vestibulum |
|
and phycobiliproteins that occur inside the thy- |
(Brett and Wetherbee, 1996). Sulfated fucose-rich |
|
lakoids of the chloroplast. The cell body is asym- |
polysaccharides can be excreted outside of the cell |
|
metric with a clearly defined dorsi-ventral/ |
(Giroldo and Vieira, 2002). |
|
right-left sides (Figs. 9.1, 9.9, 9.10). The asymmetric |
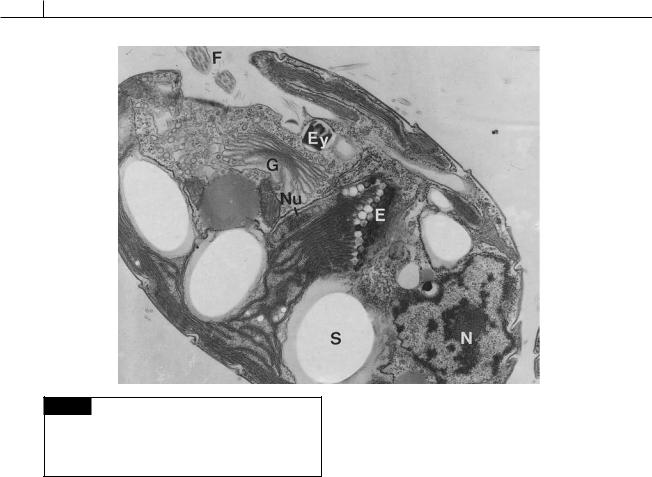
The chloroplast most likely evolved from a |
|
cell shape results in a peculiar swaying motion |
symbiosis between an organism similar to the |
|
during swimming. Most cryptophytes have a |
phagocytic cryptomonad Goniomonas and a red |
|
single lobed chloroplast with a central pyrenoid. |
alga (Kugrens and Lee, 1991; Liaud et al., 1997; |
|
|
McFadden et al., 1994). The chloroplast is sur- |
|
Cell structure |
rounded by two membranes of chloroplast endo- |
|
plasmic reticulum and the two membranes of the |
||
|
||
|
chloroplast envelope (Fig. 9.1). Between the outer |
|
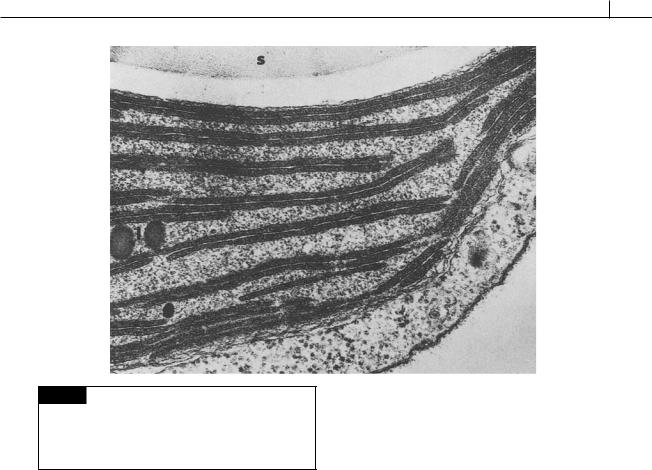
There are two apically or laterally attached |
membrane and the inner membrane of the chloro- |
|
flagella at the base of a depression. Each flagel- |
plast endoplasmic reticulum are starch grains and |
|
lum is approximately the same length as the body |
a nucleomorph (Figs. 9.1, 9.4). The nucleomorph |
|
of the cell (Figs. 9.1, 9.8, 9.9, 9.10). Depending on |
contains three minute paired-chromosomes with |
|
the species, there are one or two rows of micro- |
531 genes (humans have at least 31 000 genes) that |
|
tubular hairs attached to the flagellum. In |
encode 30 proteins targeted into the chloroplast |
|
Cryptomonas sp., the hairs on one flagellum are 2.5 |
(Douglas et al., 2001; Cavalier-Smith, 2002). The |
|
m long and in two rows whereas the hairs on the |
nucleomorph is probably the remnant of the |
|
other flagellum are only 1 m long and arranged |
nucleus of the endosymbiont in the event that led |
|
in a single row (Heath et al., 1970; Kugrens et al., |
to chloroplast E.R. The nucleomorph is sur- |
|
1987). Small, 150-nm-diameter organic scales (Fig. |
rounded by an envelope that has pores similar to |
|
9.2) are common on the flagellar surface and |
those in a nuclear envelope. The nucleomorph |
|
sometimes on the cell body (Lee and Kugrens, |
exhibits a rudimentary type of division utilizing |
|
1986). |
microtubules (Morrall and Greenwood, 1982). |
|
The outer portion of the cell, or periplast |
The nucleomorph divides in preprophase of the |
|
(Gantt, 1971), is composed of the plasma mem- |
main nucleus following basal body replication, |
|
brane and a plate, or series of plates, directly under |
but before division of the chloroplast and the |
|
the plasma membrane (Figs. 9.1, 9.10) (Kugrens |
chloroplast endoplasmic reticulum (McKerracher |