

Protistologia
.pdf418 Избранные главы общей протистологии
Вода
Встречаемость протистов определя ется наличием хотя бы небольшого ко личества свободной воды, и уже второ степенно, пресная это вода, соленая или она находится внутри тела другого
организма. Влага может просто покры вать частички почвы тонкой пленкой или поверхность листьев в виде капель росы. Протисты не способны заселять совершенно сухие биотопы. Однако многие из них образуют цисты, позво ляющие им длительное время пережи-
Рис. 349 . Население морского льда, а — слепок каналов в куске арктического льда (полу чен путем импрегнации смолой); в плавающем льду каналы заполнены гиперсоленой во д о й , б — схема распределения населяющих эти каналы о р г а н и з м о в 1 . Многоклеточные: а м ф и п о д ы , копеподы, эвфаузииды, турбеллярии; одноклеточные (на врезке): бактерии, инфузории, диатомовые, динофлагеллаты, фораминиферы (а — из: Weissenberger et al.: Limnol. Oceanogr. 37 [1992] 179; б— по разным авторам). Увел.: 15х.
1 На схеме изображено, вероятно, не арктическое, а антарктическое сообщество. Отсутствует сезонная слоистость, характерная для преобладавшего до самых последних лет в Арктике мно голетнего льда; вокруг Антрактиды же всецело доминирует однолетний лед. Планктонные фо раминиферы в обилии встречаются в антарктическом, но не в арктическом льду, зато многочис ленные в арктическом льду нематоды не указаны в составе фауны. — Прим. ред.

Экология протистов |
419 |
вать высыхание. Некоторые формируют склероции, в которых они сохраняются годами в состоянии покоя, именуемом криптобиоз. Эта стратегия делает даже пустыню средой обитания протистов, где они становятся трофонтами на ко роткий срок, после выпадения хотя бы небольшого дождя. В течение этих не скольких часов они эксцистируются, питаются, растут, пролиферируют и снова инцистируются. Протисты, реа лизующие эту стратегию, обитают не только в засушливых местообитаниях, но и в районах с умеренным климатом. Здесь их можно найти на объектах, ко торые лишь время от времени покрыты влагой: на коре деревьев, листьях, мхах и лишайниках. Слизевики и такие ин фузории, как Colpoda, сумели приспо собиться к этим нелегким условиям.
Воздушная среда, типично сухая, также используется протистами: напри мер, по воздуху распространяются ци сты или споры. Для этого некоторые слизевики (см. рис. 56, 179, 180) и ин фузория Sorogena (см. рис. 226) форми руют сорокарпы или сорусы.
При температуре ниже точки льдо образования условия тоже следует счи тать безводными, поскольку вода в твердой фазе не может служить раство рителем при метаболических процес сах. Кроме того, твердая среда не позво ляет протистам перемещаться. Неудиви тельно, что устойчивые к высыханию цисты часто также устойчивы и к вымо раживанию. Замерзание несет, по мень шей мере, две угрозы. Во-первых, сфор
мировавшиеся внутриклеточно крис таллы льда растут и разрушают мемб раны, вызывая катастрофические по вреждения вплоть до летальных. Вовторых, изменение ионного баланса ве дет к денатурации ферментов. Перед высыханием или замерзанием протисты активно выводят воду из организма. Их цисты обычно имеют сложную стенку. Считается, что именно дефицит влаги вызывает склеротизацию у слизевиков и некоторых наземных амеб (например,
Thecamoeba terricola). Предупредить формирование кристаллов льда в ци топлазме, не инцистируясь, то есть со храняя физиологическую активность, способны протисты, приспособившие ся к обитанию в концентрированных рассолах. Таковы организмы, обитаю щие в морском льду. При замерзании соленой воды в твердую фазу переходит чистая, почти без примесей Н20, поэто му по мере роста кристаллов льда идет отжим рассола. Эта жидкая фаза сохра няется в узких вертикальных каналах диаметром от 100 мкм до нескольких миллиметров (рис. 349). Увеличение концентрации солей ведет к понижению температуры замерзания. Концентрация NaCl1 в этой жидкости может достигать 144%о (при -10 °С); для сравнения, оке аническая вода имеет соленость около 38%о2. Система каналов внутри льдины населена протистами, которые не толь ко выживают в этих условиях, но и на ходят их оптимальными. Помимо диа томовых, здесь можно найти динофлагеллат, преимущественно из родов
1 Не NaCl, а сумма солей. — Прим. ред.
2 Среднее для океана значение солености составляет 35%о, а приведенная величина 38%о харак теризует некоторые краевые бассейны, например Мексиканский залив и Средиземное море, где привнос пресной воды не компенсирует испарение. — Прим. ред.

420 Избранные главы общей протистологии
Amphidinium, Gymnodinium и Glenodinium, а также некоторых хризомонад, фораминифер и инфузорий.
Чтобы выжить в ледовых кавернах, протисты изменяют состав цитоплазмы, накапливая сахара (трегалозу, маннитол, глюкозу), глицерин, аминокислоты и их дериваты (пролин, пара-аланин, таурин), метиламины и пара-диметил- сульфониопроприонат. Эти органиче ские вещества не всегда синтезируют ся самими простейшими, они могут поступать с пищей. По крайней мере, в экспериментальных условиях те гетеро трофные протисты, которых кормили толерантными к исключительно высо кой солености жгутиконосцами рода Dunaliella (Chlorophyceae), переживали глубокое замораживание и хранение в замороженном состоянии лучше, чем контрольные организмы, которых кор мили обычной пищей.
Цитоплазма гипертонична по отно шению к низкоосмотичной среде, на пример дождевой влаге, росе или клю чевой воде, поэтому вода непрерывно диффундирует в клетку. Это могло бы со временем разорвать клетку, но про тисты способны уменьшать концентра цию осмотически активных веществ, превращая их в осмотически неактив ные. Другие протисты укрепляют кле точную стенку или внутриклеточные кортикальные структуры, но большин ство удаляют избыток воды посред ством сократительных вакуолей.
Температура
Большинство протистов живет при температурах от точки замерзания воды до примерно 40 °С. Тем не менее,
встречаются и такие, которые способны выдерживать экстремальные температу ры: в термальных источниках инфузо рия Oxytricha fallax живет при темпера туре от 41 до 56 °С, а виды рода Euplo tes были найдены в морской воде при -2 °С (рис. 350).
Постепенный нагрев и затем непро должительное пребывание при 35 °С выдерживают многие виды. В природе подобные подъемы температуры случа ются из-за солнечного прогрева в таких местообитаниях, как мочажины, сфаг новая подстилка, литоральные ванны. Однако благодаря ночной прохладе (или приливу), среднесуточная темпера тура вполне терпима. С другой стороны, определенные виды имеют оптимум роста при относительно высокой темпе ратуре — например, жгутиконосцы, амебы и инфузории, населяющие мор ские и пресноводные горячие источни ки. Так, инфузория Trimyema minutus обнаружена близ Вулканического ост-
100 |
|
|
|
|
|
50 |
ч |
£ antarctious |
|
|
|
10 |
|
Е, |
vannus |
Е. |
balteatus |
|
|
|
|
|
|
• 5 |
|
|
|
/ |
|
о |
10 |
20 |
30 |
40 |
X |
Рис. 350 . Время г е н е р а ц и и различных ви дов Euplotes: Е. antarcticus (из Антарктики),
Е. vannus (из Дании), £. balteatus (из Фло риды) . Кривые показывают, что инфузории в ы д е р ж и в а ю т д о в о л ь н о б о л ь ш о й р а з м а х температур, но температура оптимального роста культуры близка к температуре сре д ы , из которой клетки были исходно выде лены (по Ли и Фенчелю) .

|
60 |
|
|
50 |
|
|
40 |
|
I |
30 |
|
О) |
|
|
о |
|
|
m |
20 |
|
в |
||
|
||
5 |
|
|
0) |
|
|
а |
10 |
|
со |
О25 30 35 40 45 50 55
Температура •
Рис. 3 5 1 . Влияние температуры на время уд воения численности инфузорий клона EV10 Trimyema minutum. Рост числа клеток в куль туре не наблюдается при температуре ниже 28 'С или выше 52 "С (из: Baumgartner et al.: J. Eukaryot. Microbiol. 49 [2002] 227).
рова (Италия) на песчаном морском дне, которое прогревается за счет под водных горячих ключей и высачиваний.
Экология протистов |
4 2 1 |
Наиболее быстрый рост инфузорий на блюдался в интервале температур от 40 до 50 °С, а кратчайший период удвое ния популяции — 6 часов — был отме чен при 48 °С (рис. 351). У некоторых свободноживущих неглерий (Schizopyrenida, Heterolobosea) максимальная скорость деления наблюдается при 40 °С
(Naegleria fowlery, N. australiensis)
(табл. 19). Эта теплолюбивость служит преадаптацией к эндопаразитизму в птицах и млекопитающих. Протист по началу приобретает способность пере живать временно в теплокровном хозя ине, а затем переходит к факультатив ному паразитизму.
Массовые скопления одноклеточ ных иногда формируются в открытом море после длительного периода тихой погоды. Помимо гетеротрофных динофлагеллат, таких как Noctiluca scintillans (рис. 352), вспышки цветения дают фототрофные гаптомонады, в ча-
Рис. 352 . Аэрофотоснимок цветения Noctiluca scintillans в Северном море (а); б — живые клетки Noctiluca (а — любезно предоставлен Дж. Восс, Киль; б — любезно предоставлен В. фон Эгмонд, Роттердам). Увел.: б — 25х.

422 Избранные главы общей протистологии
Т а б л и ц а 19. В и д ы р о д а Naegleria и мак с и м а л ь н а я т е м п е р а т у р а и х р о с т а . О п т и м а л ь н а я т е м п е р а т у р а р о с т а д л я к а ж д о г о в и д а п р и м е р н о н а 4 ° С н и ж е ( и з : d e
J o n c k h e e r e : A c t a |
Protozool . 4 1 |
[ 2 0 0 2 ] |
3 0 9 ) |
||
Виды Naegleria |
Максимальная |
|
|||
температура |
роста |
(°C) |
|||
|
|
||||
N. |
andersoni |
40 |
|
|
|
N. |
australiensis |
4 2 |
|
|
|
N. |
carteri |
45 |
|
|
|
N. |
chilensis |
30 |
|
|
|
N. |
clarki |
37 |
|
|
|
N. |
fowleri |
45 |
|
|
|
N. |
fultoni |
35 |
|
|
|
N. |
galeacystis |
35 |
|
|
|
N. |
gruberi |
39 |
|
|
|
N. |
indonesiensis |
38 |
|
|
|
N. |
italica |
4 2 |
|
|
|
N. |
jadini |
35 |
|
|
|
N. |
jamiesoni |
4 2 |
|
|
|
N. |
lovaniensis |
45 |
|
|
|
N. |
minor |
38 |
|
|
|
N. |
morganensis |
4 4 |
|
|
|
N. |
niuginensis |
45 |
|
|
|
N. |
pagei |
37 |
|
|
|
N. |
pringsheimi |
37 |
|
|
|
N. |
pussardi |
41 |
|
|
|
N. |
robinsoni |
38 |
|
|
|
N. |
sturti |
4 4 |
|
|
|
N. |
tihangensis |
4 2 |
|
|
|
Рис. 353 . Пена на побережье острова Нор- д е р н и (Германия), о б р а з о в а в ш а я с я после м а с с о в о г о цветения г а п т о м о н а д ы Phaeocystis в мае 1981 года (любезно предостав лен X. Михаелис, Нордерни) .
стности, так называемая водоросльубийца Chrysochromulina polylepis, выз вавшая «красный прилив» ранней вес ной 1988 года в Северном море. Сход ные, но не обязательно токсичные явле ния случаются по весне вдоль берегов Северного моря и Ла-Манша: в огром ном количестве развиваются разные виды гаптомонад Phaeocystis. Источае мые ими углеводы производят пену высотой до метра (рис. 353).
Кислород
Присутствие растворенного кисло рода в окружающей жидкости необхо димо для большинства протистов. Од нако они гораздо менее, чем много клеточные, требовательны к концент рации кислорода, т.к. при их малых размерах диффузия гораздо более эф фективна. Многие виды населяют мес тообитания с крайне низким содержа нием кислорода, а некоторые способ ны существовать даже при полном его отсутствии, в сильно восстановитель ной среде, например в активном иле очистных сооружений. В таких ана эробных местообитаниях обнаружива ются инфузории родов Metopus и Саеnomorpha или свободноживущие дипломонады родов Trepomonas и Нехаmita. Многие из обитающих в анаэ робных условиях протистов содержат в цитоплазме метаногенных бактерий, которые утилизируют Н2, образующий ся у хозяина в качестве конечного продукта метаболизма (рис. 354). В то же время, в таких биотопах обитают многие протисты, например дипломонады, которые полностью лишены митохондрий и иных окислительно-
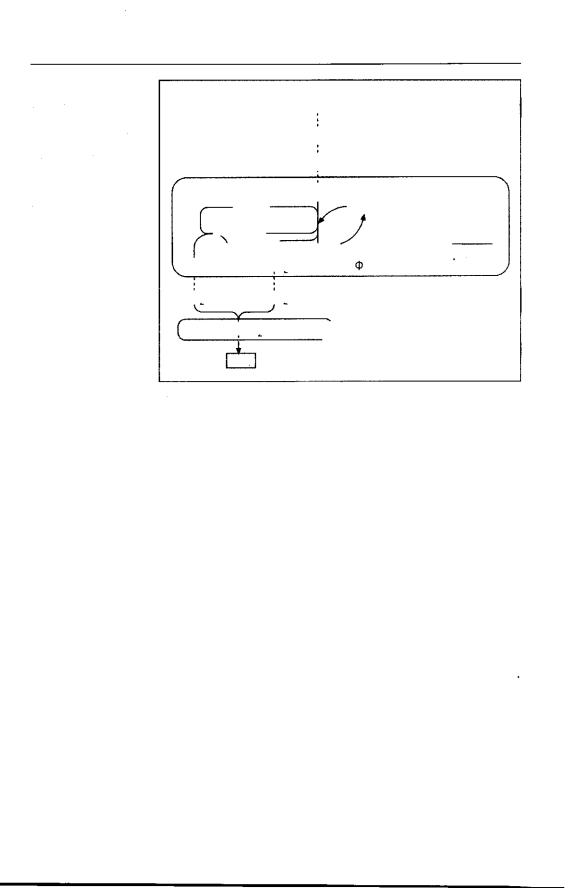
Экология протистов |
4 2 3 |
-2 F d + -
|
|
^__2 F d - |
||
|
|
2 Н + |
Г |
|
|
|
ft |
С О , |
|
Рис . |
3 5 4 . С х е м а пу |
|
||
|
|
|||
тей |
в з а и м о д е й с т в и я |
4 Н , |
С О , |
|
м е ж д у г и д р о г е н о с о - |
||||
|
|
|||
м а м и и э н д о б и о н т - |
|
|
||
ными метаногенными |
N- Н,0 |
|||
б а к т е р и я м и в а н а э |
|
|
||
р о б н ы х и н ф у з о р и я х |
сн |
4 |
||
(по Важинеру). |
||||
|
|
|||
Углеводы
(Гликолиз) Цитоплазма инфузории
Пир'уват
Пир'уват Гидрогеносома •КоА
А ц е т и п - К о А - т — ^ З Д : ^ А ц е т а т !
АДФ АТФ
J Метаногенная бактерия
восстановительных органелл (то есть, гидрогеносом и гликосом. — Прим. ред.).
Примечательно, что многие инфузо рии, обитающие в стоячих водоемах или в среде с крайне низкой концентра цией кислорода, содержат в цитоплаз ме одноклеточные зеленые водоросли
Chlorella и Trebouxia, обобщенно назы ваемые зоохлореллами. Эти эндобионты могут присутствовать облигатно, как у Climacostomum virens или Paramecium bursaria, или факультативно, как у ряда видов Vorticella или Frontonia. Выгода цилиат в том, что фотосинтетически активные зоохлореллы обеспечивают их сахарами и, вероятно, кислородом.
Инфузории, узко адаптированные к низкой концентрации кислорода, в озе рах с сезонно развивающейся аноксией мигрируют вслед за кислородно-бескис лородной границей. Эти организмы,
особенно Loxodes и Spirostomum, прово дят большую часть времени в осадке, но вслед за установлением стратифика ции водного столба и развитием аноксии в придонных горизонтах они всплывают к границе между аэрирован ным и бескислородным слоями воды. Таким образом, популяция совершает вертикальную сезонную миграцию ам плитудой в несколько метров (рис. 355). Гравитактическая ориентация у Loxodes осуществляется, вероятно, при участии специальной органеллы — тельца Мюллера (см. рис. 116, 117).
Субстраты
Наличие субстрата и его физичес кие и химические свойства существен ны для сидячих и передвигающихся по поверхности организмов. С физической точки зрения, важна низкая скорость

424 Избранные главы общей протистологии
Рис. 355. Вертикальное распределение популяции Loxodes в эвтрофном озере (Esthwaite Water, Lake District, England) в зависимости от концентрации 02 в придонном слое воды. В пе риод летней с т р а т и ф и ц и р о в а н н о е ™ водного столба (июнь - сентябрь) зона дефицита кис лорода ( 0 - 2 г/л) захватывает толщу воды 5 - 6 метров надо д н о м . Значительная часть попу ляции Loxodes летом мигрирует вверх вслед за границей дизаэробной зоны, хотя опреде ленная доля особей периодически уходит в анаэробный гиполимнион . Когда стратифика ция водного столба нарушается (начало октября), все Loxodes возвращаются в бентос (из: Finlay and Fenchel: Freshwater Biological Association Annual Report 54 [1986] 73).
потока воды, которая наблюдается вбли зи поверхности субстрата. Поэтому пе риферические участки водотоков и за стойные зоны рек обычно густо заселе ны протистми.
Сидячие формы составляют суще ственную долю сообщества протистов почти в каждом биотопе. Численность многих свободноподвижных видов бы вает особенно велика в непосредствен-

Поведение протистов |
4 2 5 |
Рис. 356. Схема фагоцитоза у Amoeba proteus. ж — жертва (по Хаберей и Стокему).
ной близости от частиц осадка и агре гатов детрита, так как здесь обильны мелкие пищевые объекты и растворен ные питательные вещества.
Фильтраторы неподвижно прикреп ляются, чтобы трансформировать пла вательное биение ресничек в движение потока, приносящего пищевые частицы. В то же время, чтобы иметь возмож ность создавать эффективные токи жид кости, клетки должны быть приподня ты над субстратом. Обеспечить это по зволяют стебельки и удлиненная форма тела, как, например, у инфузорий Vorticella и Stentor и хоанофлагеллат. Проти сты повышают эффективность фильтра ции, формируя плавающие колонии; в качестве примера можно привести ша ровидные колонии хоанофлагеллат и небольшие колонии таких инфузорий, как Ophrydium versatile.
Ползущая амеба улавливает жертву, обтекая ее прижатыми к субстрату лобоподиями (рис. 356). Плазмодий Reticulomyxa (Foraminifera) растягивает над поверхностью субстрата обширную сеть ретикулоподий и ловит весьма крупную добычу, в частности, коловра ток и колонии Volvox (рис. 357).
Некоторые сидячие инфузории предпочитают прикрепляться к органи ческим или живым субстратам (рис. 358). Особенно это относится к пред ставителям рода Spirochona, которые обнаруживаются исключительно на жа берных пластинках гаммарид; а еще это характерно для ряда перитрих, которые
Рис. 357 . Участок сети ретикулоподий Reticulomyxa filosa, которые, как показывает от б р а с ы в а е м а я и м и тень, м о г у т п р и п о д н и маться над субстратом . Увел.: 200х.
Рис. 3 5 8 . К р у г о р е с н и ч н ы е и н ф у з о р и и н а антеннах листоногого рачка (из: Hausmann and Rambow: M i k r o k o s m o s 7 4 [ 1 9 8 5 ] 208) . Увел.: 110x.

426 Избранные главы общей протистологии
прикрепляются к ракообразным. Мно гие суктории видоспецифичны по отно шению к животным и растениям, на которых они обитают. Например, Tokophrya lemnae селится только на ри зоидах обычной ряски (Lemna minor). Эволюционное значение такой крайней специализации, возможно, состоит в избегании конкуренции. Механизмы распознавания субстрата у протистов неизвестны.
Биотические факторы
Как уже упоминалось, самыми важ ными биотическими факторами являют ся пищевые ресурсы, конкуренция и взаимоотношения хищника и жертвы.
Такие организмы, как фораминиферы, с периодом воспроизводства не сколько недель или даже месяцев, вы живают только в очень стабильных ус ловиях. Здесь они в типичном случае представляют одно постоянное звено в довольно сложной структуре биоцено за. Наоборот, протисты с репродуктив ным циклом лишь в несколько часов истощают ресурсы очень быстро. Попу ляции амеб, жгутиконосцев или инфу зорий, как правило, исчезают столь же быстро, как и появились.
Первая ступень формирования био ценоза — заселение нового биотопа. На искусственном субстрате, размещенном в пресных прудах, в течение трех не дель эксперимента поселилось около 60 видов протистов. С другой стороны, в особых случаях колонизация пойдет очень медленно, если, например, новое местообитание находится на значитель ном расстоянии от источника вселяю щихся видов. Иллюстрацией может слу
жить заселение вулканического остро ва Суртсей, возникшего южнее Ислан дии в 1963 году: за двухлетний период в экспонированных емкостях с пресной водой было обнаружено не более 15 видов протистов.
Пионерными видами обычно высту пают жгутиконосцы, в частности, воротничковые жгутиконосцы и бодониды, которые затем сменяются мелкими амебами и инфузориями. Пищей им служат бактерии, цианобактерии и од ноклеточные водоросли, в частности,
|
|
|
|
Euplotes |
patella |
|
|
|
|
|
Суктории |
|
|
|
|
|
|
Halteria grandinella |
||
|
|
|
|
Chilodonella |
cucullus |
|
|
|
|
|
Coleps hirtus |
||
|
|
|
|
Microthorax |
pusillus |
|
|
|
|
|
Litonotus |
lamella |
|
|
|
|
|
Paramecium |
caudatum |
|
|
|
|
|
Vorticella |
|
|
|
|
|
|
Stylonychia |
putrina |
|
|
|
|
|
Ankistrodesmus |
||
|
|
|
|
Colpidium |
campylum |
|
|
|
|
|
Chilomonas |
|
|
|
|
|
|
Scenedesmus |
||
|
|
|
|
Cyclidium |
citrullus |
|
|
|
|
|
Glaucoma |
scintillans |
|
|
|
|
|
Euglena |
|
|
|
|
|
|
Диатомовые |
|
|
|
|
|
|
Амебы |
|
|
|
|
|
|
Гетеротрофные |
||
|
10 |
20 |
30, |
жгутиконосцы |
||
|
|
|
|
|||
Рис. 359 . Сукцессия п р о т и с т о в |
из п р о б ы , |
|||||
взятой |
в е с т е с т в е н н о м м е с т о о б и т а н и и и |
|||||
затем |
в ы р а щ е н н о й в л а б о р а т о р и и на обо |
|||||
гащенной целлюлозой среде . Эксперимент длился 35 дней (по Бику).

диатомовые. Пищедобывающая актив ность протистов во многом определяет качественный и количественный состав популяций жертв. В ходе становления таких биоценозов среда преобразуется, и формируются новые экологические ниши, пригодные для заполнения, в первую очередь, хищными протистами.
Последовательность появления ви дов в сообществе называется сукцесси ей. Лабораторные эксперименты пока зали, что сообщества протистов неста бильны и что они претерпевают карди нальную перестройку очень быстро, часто в течение дня (рис. 359). Такой ход сукцессии, вероятно, отражает ис тощение определенных пищевых ресур сов или смену доминирующих хищни ков. Влияние хищника на популяцию бактериотрофной инфузории легко про демонстрировать на простой пищевой цепи из трех видов (рис. 360). Если в растущую культуру Vorticella добавить хищную инфузорию Hemiophrys, коли чество вортицелл резко упадет, тогда как численность Hemiophrys достигнет максимума. После истощения популя ции жертвы численность хищника рез-
tSOr
Экология протистов |
4 2 7 |
ко снизится. Вортицеллы снова размно жатся, питаясь обильно разросшимися бактериями. Цикл начнется снова. Сход ные сукцессии по типу хищник—жерт ва известны для пар инфузорий Didi nium nasutum и Paramecium caudatum, а также Trachelius ovum и Ophrydium versatile.
В естественных биотопах ситуация более сложна из-за влияния дополни тельных факторов на плотность популя ций. Один из таких факторов — вклю чение в пищевую цепь многоклеточных хищников. Некоторые протисты будут съедены, к примеру, личинками рыб, улитками, плоскими червями или олигохетами; другие погибнут, не выдер жав конкуренции за пищу с коловратка ми (рис. 361).
Протисты способны заселять биото пы, где очень мало или совсем нет мно гоклеточных. Как показано на примере морской интерстициали, микробиотопы могут быть слишком малы для проник новения в них многоклеточных живот ных. Другой пример — это биотопы с экстремальными физическими или хи мическими характеристиками, такие
в р е м я ( ч )
Рис. 360 . Эксперимент по популяционной динамике в трехкомпонентной системе: бакте рия, инфузория Vorticella и инфузория Hemiophrys (по Курдсу).