

-
Биосфера. Учение в.И. Вернадского о биосфере. Ноосфера. Термин «биосфера» впервые применил австрийский геолог э. Зюсс (1873), определяя им пространство органической жизни на Земле.
Выдающаяся роль в развитии учения о биосфере принадлежит русскому ученому В.И. Вернадскому. В своем классическом труде «Биосфера» (1926 г.) он по существу переоткрыл это понятие, придав ему смысл и статус глобальной системы, в которой все живые организмы так взаимодействуют между собой и с окружающей средой, что оказывают определяющее влияние на планетарные геохимические и энергетические превращения.
Биосфера – это часть земного шара, в пределах которой существует жизнь. Она представляет собой оболочку Земли, состоящую из атмосферы, гидросферы и верхней части литосферы, которые взаимно связаны сложными биохимическими циклами миграции вещества и энергии.
Выдающийся русский учёный В.И. Вернадский (1868-1945), один из создателей современного взгляда на биосферу, определил её как наружную оболочку Земли, область распространения жизни, основа которой - взаимодействие живого и косного вещества. Согласно современным представлениям, биосфера – это своеобразная оболочка Земли, содержащая всю совокупность живых организмов и ту часть вещества планеты, которая находится в непрерывном обмене с этими организмами. В очень сложной по составу, строению и организованности биосфере можно выделить сравнительно небольшое число основных компонентов:
- живое вещество – это совокупность тел живых организмов, населяющих Землю вне зависимости от их систематической принадлежности. Общий вес живого вещества оценивается в 2400 млрд. т (в сухом виде). По определению В.И Вернадского, живое вещество является одной из самых могущественных геохимических сил нашей планеты;
- биогенное вещество – это химические соединения, возникшие в результате жизнедеятельности организмов. При этом образующиеся химические соединения могут и не входить в состав тел организмов (каменный уголь, торф, гумус, мел, известняк, нефть и т. д.);
- косное вещество – это вещество, которое образуется без участия живого вещества (горные породы земной коры неорганического происхождения, вода, компоненты воздуха);
- биокосное вещество – это продукты переработки горных и осадочных пород организмами;
- вещество космического происхождения. Биосфера в главных своих чертах может быть охарактеризована по отдельным оболочкам, которые она охватывает: нижняя часть атмосферы, гидросфера и верхняя часть литосферы. Атмосфера – это газовая оболочка, окружающая Землю. Атмосфера имеет слоистую структуру: тропосфера (до 10 км), стратосфера (10-47 км), мезосфера (47-80 км), термосфера (80-1000 км). На высотах свыше 1000 км находится экзосфера. В состав атмосферы входят: N (78,08 % по объёму), O2 (20,95 %), Ar (0,93 %), CO2 (0,03 %), Ne, He, CH4, Kr, H2, Xe, водяной пар, O3 и др. Одним из важнейших компонентов атмосферы является озон О3. Толщина озонового слоя 2,3-5,2 мм. Гидросфера – водная оболочка Земли. Гидросфера находится в тесной взаимосвязи с литосферой (подземные воды), атмосферой (парообразная влага) и живым веществом биосферы, в которое она входит в качестве обязательного компонента. Подавляющая часть природных вод слагает Мировой океан (94 %), который представляет собой уникальную природную систему биосферы. В ней происходит грандиозный процесс обмена и трансформации энергии и вещества нашей планеты. Вода является самым мощным поглотителем солнечного тепла на поверхности Земли, поэтому решающая роль в поглощении солнечной энергии на нашей планете принадлежит Мировому океану. Литосфера – это верхняя твёрдая оболочка Земли. В состав биосферы входят верхние горизонты литосферы (литобиосфера), подвергающиеся ныне или подвергавшиеся в прошлом воздействию живых организмов. Общий химический состав литобиосферы определяют немногие элементы (О, Si, Al, Fe, Са, Mg, Na, К), которые слагают основную её массу. При этом наиболее распространённым элементом является кислород, который прочно связан химически с другими элементами. В литосферу живые организмы проникают на ничтожную глубину. Основная их масса сосредоточена в верхнем слое - почве мощностью в несколько десятков сантиметров. Ноосфера — предположительно новая, высшая стадия эволюции биосферы, становление которой связано с развитием общества, оказывающего глубокое воздействие на природные процессы. Согласно В. И. Вернадскому, «в биосфере существует великая геологическая, быть может, космическая сила, планетное действие которой обычно не принимается во внимание в представлениях о космосе… Эта сила есть разум человека, устремленная и организованная воля его как существа общественного».
-
Законы и следствия пищевых отношений. Отношения в системе «хищник-жертва». Пищевые отношения не только обеспечивают энергетические потребности организмов. Они играют в природе и другую важную роль — удерживают виды в сообществах, регулируют их численность и влияют на ход эволюции. Пищевые связи чрезвычайно разнообразны.
Типичные хищники. У них развито специальное охотничье поведение. Обычно это сильные и активные животные.
Паразиты живут в условиях избытка пищи, которую не надо активно добывать, и используют хозяев как среду своего обитания. У них упрощено строение и ослаблены связи с внешним миром.
Животные-собиратели тратят энергию на поиск семян или насекомых, т. е. мелкой добычи. У них развита поисковая активность, но нет охотничьего поведения.
Пасущиеся виды не тратят много сил на поиск корма, обычно его достаточно много вокруг, и основное время у них уходит на поглощение и переваривание пищи.
В водной среде широко распространен такой способ овладения пищей, как фильтрация, на дне — заглатывание и пропускание через кишечник грунта вместе с пищевыми частицами.
Последствия пищевых связей наиболее ярко проявляются в отношениях хищник — жертва.
Если хищник питается крупными, активными жертвами, которые могут убегать, сопротивляться, прятаться, то в живых остаются те из них, кто делает это лучше других, т. е. имеет более зоркие глаза, чуткие уши, развитую нервную систему, мускульную силу. Таким образом, хищник ведет отбор на совершенствование жертв, уничтожая больных и слабых. В свою очередь, и среди хищников тоже идет отбор на силу, ловкость и выносливость. Эволюционное следствие этих отношений — прогрессивное развитие обоих взаимодействующих видов: и хищника, и жертвы.
Если же хищники питаются малоактивными либо мелкими, не способными сопротивляться им видами, это приводит к другому эволюционному результату. Погибают те особи, которых хищник успевает заметить. Выигрывают менее заметные или чем-то неудобные для захвата жертвы. Так осуществляется естественный отбор на покровительственную окраску, твердые раковины, защитные шипы и иглы и другие орудия спасения от врагов.
Самый существенный результат трофических взаимосвязей — сдерживание роста численности видов.
Для каждой пары видов хищника и жертвы результат их взаимодействия зависит прежде всего от их количественных соотношений. Если хищники ловят и уничтожают своих жертв примерно с той же скоростью, с какой эти жертвы размножаются, то они могут сдерживать рост их численности (устойчивое природное сообщество). Если скорость размножения жертв выше, чем скорость поедания их хищниками, происходит вспышка численности вида. Обратный результат — полное уничтожение жертвы хищником — в природе очень редок (с падением численности какого-либо вида жертв в природе хищники переключаются на другую, более доступную добычу).
Правило Гаузе (Правило конкурентного исключения): Два вида с близкими экологическими требованиями не могут длительное время существовать совместно. Правила Тинемана: - чем больше видов живёт в одном месте, тем более детально они отличаются друг от друга по использованию сходных ресурсов; - чем беднее видами сообщество, тем выше может быть численность каждого отдельного вида. Экологическая ниша – положение, которое занимает вид в сообществе. Гаузе в опыте с инфузорией туфелькой и дидениумом было доказано, что взаимодействия хищника и жертвы могут при известных условиях приводить к регулярным циклическим колебаниям численности обоих видов. Ход этих циклов можно рассчитать и предсказать, зная некоторые исходные количественные характеристики видов. Количественные законы взаимодействия видов в их пищевых связях очень важны для практики.
Численность
В результате перепромысла, по вине людей уже исчез с лица Земли ряд видов, бывших когда-то очень многочисленными: американские бизоны, европейские туры, странствующие голуби и другие.
При случайном или намеренном уничтожении хищников какого- либо вида сначала возникают вспышки численности его жертв. Это тоже приводит к экологической катастрофе либо в результате подрыва видом собственной кормовой базы, либо — распространения инфекционных заболеваний, которые часто бывают гораздо более губительны, чем деятельность хищников. Возникает явление экологического бумеранга, когда результаты оказываются прямо противоположными начальному направлению воздействия. Поэтому грамотное использование природных экологических законов — основной путь взаимодействия человека с природой.
Математическая модель: Модель Лотки — Вольтерра. Не правильное использование математической модели привело в 1930-1988 гг. к истреблению трески и мойвы.
-
Круговорот фосфора. Этот элемент входит в состав генов и молекул, переносящих энергию внутрь клеток. В различных минералах фосфор содержится в виде неорганического фосфат-иона (РО43-). Фосфаты растворимы в воде, но не летучи. Растения поглощают фосфат-ион из водного раствора и включают фосфор в состав различных органических соединений, где он выступает в форме так называемого органического фосфата. По пищевым цепям фосфор переходит от растений ко всем прочим организмам экосистемы. При каждом переходе велика вероятность окисления содержащего фосфор соединения в процессе клеточного дыхания для получения организмом энергии. Когда это происходит, фосфат в составе мочи или её аналога вновь поступает в окружающую среду, после чего снова может поглощаться растениями и начинать новый цикл.
У фосфора нет газовой фазы и, следовательно, нет «свободного возврата» в атмосферу. Попадая в водоёмы, фосфор насыщает, а иногда и перенасыщает экосистемы. Обратного пути, по сути дела, нет. Что-то может вернуться на сушу с помощью рыбоядных птиц, но это очень небольшая часть общего количества, оказывающаяся к тому же вблизи побережья. Океанические отложения фосфата со временем поднимаются над поверхностью воды в результате геологических процессов, но это происходит в течение миллионов лет.
Следовательно, фосфат и другие минеральные биогены почвы циркулируют в экосистеме лишь в том случае, если содержащие их отходы жизнедеятельности откладываются в местах поглощения данного элемента. В естественных экосистемах так в основном и происходит. Когда же в их функционирование вмешивается человек, он нарушает естественный круговорот, перевозя, например, урожай вместе с накопленными из почвы биогенами на большие расстояния к потребителям.
-
Круговорот азота. Основная часть атомов азота находится в воздухе, который на 78 % состоит из одноимённого газа (N2). Однако растения не могут усваивать его непосредственно; для этого азот должен входить в состав ионов аммония (NH4+) или нитрата (NH3-). К счастью, некоторые бактерии и ряд сине-зелёных водорослей способны превращать газообразный азот в аммонийную форму в ходе так называемой азотфиксации. Важнейшую роль среди азотфиксирующих организмов играют бактерии, живущие в клубеньках на корнях бобовых растений. По пищевым цепям органический азот передаётся от бобовых другим организмам экосистемы.
Когда в процессе клеточного дыхания белки и другие содержащие азот органические соединения расщепляются, азот выделяется в среду главным образом в аммонийной форме. Некоторые бактерии могут переводить ее в нитратную форму. Важно то, что обе эти формы могут усваиваться любыми растениями. В результате азот совершает круговорот как минеральный биоген. Однако такая минерализация обратима, поскольку другие почвенные бактерии постепенно превращают нитраты снова в газообразный азот. Правда, часть его окисляется в воздухе во время грозовых разрядов и поступает в почву с дождевой водой, но таким способом его фиксируется в 10 раз меньше, чем с помощью бактерий.
Таким образом, все естественные экосистемы зависят от азотфиксирующих организмов, поэтому крайне важна роль бактерий в клубеньках бобовых растений. Это семейство включает огромное число представителей – от клевера до тропических деревьев и пустынных кустарников. В каждой крупной наземной экосистеме -–от дождевых экваториальных лесов до тундры – есть характерные для неё виды бобовых. Интересно отметить, что бобовые обычно первыми заселяют гари – на них процесс реколонизации идёт значительно медленнее из-за недостатка в почве доступного азота. В водных экосистемах круговорот азота выглядит сходным образом, но здесь в роли основных азотфиксаторов выступают сине-зелёные водоросли.
Люди научились создавать искусственные экосистемы, выращивая урожаи кукурузы, пшеницы и других зерновых культур без участия бобовых. При этом азот воздуха фиксируется на химических заводах. Искусственно полученные аммоний и нитрат представляют собой основные ингредиенты минеральных удобрений. Однако их высокая цена вынуждает специалистов реконструировать естественные условия, чередуя в севообороте бобовые и остальные культуры.
-
Свойства биосферы как глобальной биологической системы. Биосфера – система с прямыми и обратными (отрицательными и положительными) связями, которые, в конечном счёте, обеспечивают механизмы её функционирования и устойчивости.
Биосфера – централизованная система. Центральным звеном её выступают живые организмы (живое вещество). Это свойство всесторонне раскрыто В.И.Вернадским, но, к сожалению, часто недооценивается и в настоящее время: в центр биосферы или её звеньев ставится только один вид – человек.
Биосфера – открытая система. Её существование немыслимо без поступления энергии извне. Она испытывает воздействие космических сил, прежде всего солнечной активности.
Биосфера – саморегулирующаяся система, для которой, как отмечал В.И.Вернадский, характерна организованность. В настоящее время это свойство называют гомеостазом, понимая под ним способность возвращаться в исходное состояние, гасить возникающие возмущения включением ряда механизмов. Гомеостатические механизмы связаны в основном с живым веществом. Биосфера за свою историю пережила ряд таких возмущений, многие из которых были значительными по масштабам, но справлялась с ними благодаря действию гомеостатических механизмов и, в частности, принципа, который в настоящее время носит название Ле Шателье-Брауна: при действии на систему сил, выводящих её из состояния устойчивого равновесия, последнее смещается в том направлении, при котором эффект этого воздействия ослабляется.
Опасность современной экологической ситуации связана прежде всего с тем, что нарушаются многие механизмы гомеостаза и принцип Ле Шателье-Брауна если не в планетарном, то в региональных масштабах. Их следствие – региональные кризисы. Результатом этого является либо распад экосистем (например, расширяющиеся площади опустыненых земель), либо появление неустойчивых, практически лишённых свойств гомеостаза систем типа агроценозов или урбанизированных (городских) комплексов.
Биосфера – система, характеризующаяся большим разнообразием. Последнее обусловливается многими причинами и факторами. Это и разные среды жизни; и разнообразие природных зон, различающихся по климатическим, гидрологическим, почвенным, биотическим и другим свойствам; и наличие регионов, различающихся по химическому составу (геохимические провинции); и, самое главное, объединение в рамках биосферы большого количества элементарных экосистем со свойственным им видовым разнообразием.
Разнообразие биосферы за счёт элементарных экосистем по вертикали обусловливается ярусностью или экогоризонтами растительного покрова и связанных с ними животных организмов, а в горизонтальном направлении неравномерностью распределения организмов и их группировок и связанных с ними факторов.
Для любой природной системы разнообразие – одно из важнейших её свойств. Это свойство настолько универсально, что сформулировано в качестве закона (автор его У.Р.Эшби).
Важное свойство биосферы – наличие в ней механизмов, обеспечивающих круговорот веществ и связанную с ним неисчерпаемость отдельных химических элементов и их соединений. Только благодаря круговоротам обеспечивается непрерывность процессов в биосфере.
Важно подчеркнуть, что, в конечном счете, и планетарные проблемы будут решаться тем успешнее, чем конкретнее становятся и решаются проблемы локального плана. Такая постановка задач в конкретном виде достижима на уровне элементарных единиц биосферы – экосистем или биогеоценозов.
-
Особенности зависимости организма от среды на разных этапах онтогенеза. Критические периоды развития. Учение о критических периодах развития было создано в 1921 г. К. Стоккар-дом и в дальнейшем значительно углублено и расширено П. Г. Светловым. Индивидуальное развитие, по воззрениям П. Г. Светлова, состоит из небольшого числа этапов, каждый из которых начинается критическим периодом, за которым следуют этапы видимой дифференциации и роста. Критические периоды характеризуются наиболее высокой чувствительностью к воздействиям вредных факторов внешней среды. В ранних стадиях эмбрионального развития критические периоды относятся к развитию всего организма, позднее отрицательное влияние определенных факторов сказывается на формировании отдельных органов — тех, которые в данный момент претерпевают наиболее активные формообразовательные процессы. Внешние факторы, к которым организм (или отдельный орган) весьма чувствителен в определенные периоды, могут существенным образом влиять на его развитие. Различные воздействия в один и тот же период могут вызывать сходные отклонения. И наоборот, один и тот же фактор, действующий на разных этапах, вызывает различные изменения, т. е. тип аномалии в значительной степени зависит от стадии развития, во время которой на организм оказал действие тератогенный агент.
Биологический смысл повышения чувствительности к внешним воздействиям в критические периоды заключается в обеспечении восприятия зародышем и его частями сигналов, ответом на которые являются определенные процессы индивидуального развития.
Наиболее высокой чувствительностью к повреждающим агентам обладают зародыши во время имплантации (первый критический период), соответствующий 7—8-му дню эмбриогенеза, и во время плацентации (второй критический период). Плацентация приходится на 3—8-ю неделю эмбриогенеза и совпадает с этапом формирования зачатков органов.
Повреждающие факторы внешней среды (химические агенты, в том числе лекарственные, радиация и др.) могут оказывать неодинаковое влияние на зародыши, находящиеся в разных стадиях развития: эмбриотоксическое или тератогенное. Эмбриотоксическое действие повреждающих факторов характерно для первого критического периода, тератогенное — для второго.
В период имплантации зародыш либо погибает (при повреждении многих бластомеров), либо дальнейший эмбриональный цикл не нарушается (при сохранности большого числа бластомеров, способных к полипотентному развитию). При поражении зародыша в период плацентации и органогенеза характерно возникновение уродств. При этом пороки развития образуются в тех органах, которые в момент действия повреждающих агентов находились в процессе активной дифференцировки и развития. У различных органов эти периоды не совпадают во времени. Поэтому при кратковременном действии тератогенного фактора формируются отдельные аномалии развития, при длительном — множественные.
Согласно учению о двух критических периодах эмбриогенеза, для снижения частоты гибели зародышей и врожденных пороков развития необходимо охранять организм женщины от неблагоприятных воздействий окружающей среды именно в первые 3—8 нед. беременности. Хотя дальнейшие исследования доказали, что по отношению к ряду повреждающих агентов эмбрион и плод человека обладают высокой чувствительностью и после завершения плацентации и активного органогенеза. К критическим периодам фетального развития относят 15—20-ю недели беременности (усиленный рост головного мозга) и 20—24-ю недели (формирование основных функциональных систем организма).
-
Антропогенные экосистемы. Отличие от естественных экосистем. Среди антропогенных экосистем выделяют два основных вида – агроэкосистемы (сельскохозяйственные экосистемы, агроценозы) и урбосистемы (урбанистические системы).
Агроэкосистемы - искусственные экосистемы, которые возникают в результате сельскохозяйственной деятельности человека для получения продукции автотрофов (урожая). В агроэкосистемах так же, как и в естественных сообществах, имеются продуценты (культурные растения и сорняки), консументы (насекомые, птицы, мыши и т.д.) и редуценты (грибы и бактерии). Обязательным звеном пищевых цепей в агроэкосистемах является человек. Основными отличиями агроценозов от естественных биоценозов являются:
- незначительное видовое разнообразие;
- короткие цепи питания;
- неполный круговорот веществ (часть питательных элементов выносится с урожаем);
- источник энергии – Солнце и деятельность человека;
- «эволюция» за счет искусственного отбора, производимого человеком;
- отсутствие саморегуляции и существования без поддержки человека.
Таким образом, агроценозы – это неустойчивые системы, способные существовать только при поддержке человека.
Урбосистемы (урбанистические системы) - искусственные системы, которые возникают в результате развития городов и представляющие собой средоточие населения, жилых зданий, промышленных, бытовых, культурных объектов и т.д. В составе урбоэкосистем выделяют: промышленные зоны, включающие промышленные объекты различных отраслей хозяйства; селитебные зоны с жилыми домами, административными зданиями, объектами быта, культуры и т.п.; рекреационные зоны (лесопарковые), предназначенные для отдыха людей (лесопарки, базы отдыха и т.д.)
Кроме того, значительное место в инфраструктуре городов занимают - транспортные системы и сооружения (автомогистрали, железные дороги, заправочные станции, гаражи, метрополитен и т.п.).
Существование урбоэкосистем зависит от энергии горючих ископаемых, атомной промышленности и агроэкосистем. Сравнение природной и упрощенной антропогенной экосистем (по Миллеру, 1993):
|
Природная экосистема |
Антропогенная экосистема |
|
Получает преобразует, накапливает солнечную энергию |
Потребляет энергию ископаемого и ядерного топлива. |
|
Продуцирует кислород и потребляет диоксид углерода. |
Потребляет кислород и продуцирует диоксид углерода при сгорании ископаемого топлива . |
|
Формирует плодородную почву. |
Истощает или представляет угрозу для плодородных почв. |
|
Накапливает, очищает и постепенно расходует воду. |
Расходует много воды, загрязняет ее. |
|
Создает местообитания различных видов дикой природы. |
Разрушает местообитания многих видов дикой природы. |
|
Бесплатно фильтрует и обеззараживает загрязнители и отходы |
Производит загрязнители и отходы, которые должны обеззараживаться на счет населения. |
|
Обладает способностью самосохранения и самовосстановления. |
Требует больших затрат для постоянного поддержания и восстановления. |
-
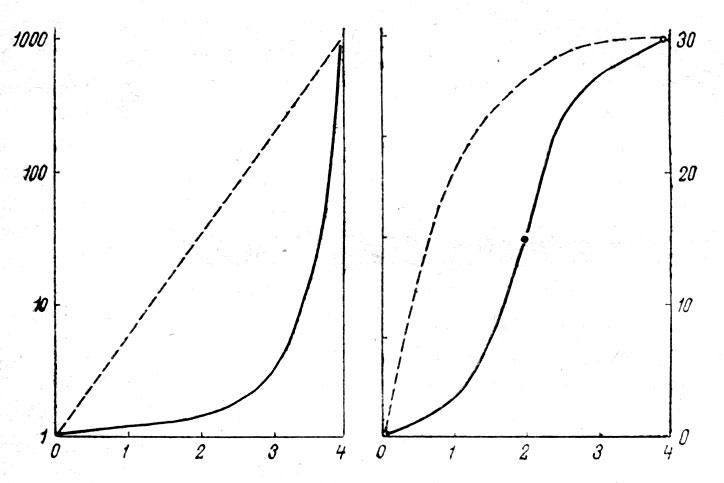
Смертность как популяционный показатель. Кривые выживания. Рождаемость - это способность популяции к увеличению, или число потомков, производимых одной самкой за 1 год. Максимальная рождаемость - теоретически максимально возможное количество особей, образующихся в идеальных условиях при отсутствии лимитирующих факторов, и размножение ограничивается лишь физиологическими факторами. Экологическая, или реализуемая рождаемость - появление новых особей при фактических условиях среды. Смертность - гибель особей за единицу времени в отсутствие лимитирующих факторов. Экологическая, или реализуемая смертность - гибель особей за единицу времени при фактических условиях среды. Разность между рождаемостью и смертностью есть некий результирующий параметр, который определяет реальную динамику численности у данной популяции. По мере роста популяции происходит снижение доступных каждой особи ресурсов среды. При истощении ресурсов рост популяции тормозится и, в конце концов, прекращается. Смертность, как и рождаемость, сильно варьируют с возрастом. Поэтому определяют экологическую смертность для различных экологических групп и вычерчивают кривые выживания.
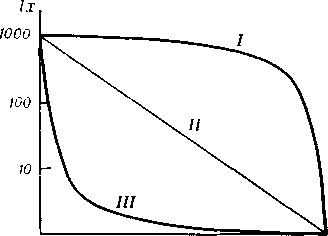 Кривая
выживания -
график, показывающий, как по мере старения
снижается численность особей одного
возраста в популяции. Различают 3 осн.
типа К. в., между которыми возможны все
переходы. Кривая
I соответствует
популяции, большинство членов которой
доживает до возраста, близкого к
максимально возможному для данного
вида; характерна для популяций некоторых
крупных млекопитающих. Кривая
II отражает
равную вероятность гибели особей в
любом возрасте и свойственна мн. природным
популяциям птиц и пресмыкающихся. Кривая
III соответствует
очень высокой смертности в раннем
возрасте, а для особей, переживших этот
период, вероятность смерти низка. Такая
кривая характерна для мн. растений,
беспозвоночных и рыб, у которых осн.
гибель приходится на ранние стадии
развития. Данные, служащие для построения
К. в., используются также в демографических
таблицах (таблицах
выживания).
Кривая
выживания -
график, показывающий, как по мере старения
снижается численность особей одного
возраста в популяции. Различают 3 осн.
типа К. в., между которыми возможны все
переходы. Кривая
I соответствует
популяции, большинство членов которой
доживает до возраста, близкого к
максимально возможному для данного
вида; характерна для популяций некоторых
крупных млекопитающих. Кривая
II отражает
равную вероятность гибели особей в
любом возрасте и свойственна мн. природным
популяциям птиц и пресмыкающихся. Кривая
III соответствует
очень высокой смертности в раннем
возрасте, а для особей, переживших этот
период, вероятность смерти низка. Такая
кривая характерна для мн. растений,
беспозвоночных и рыб, у которых осн.
гибель приходится на ранние стадии
развития. Данные, служащие для построения
К. в., используются также в демографических
таблицах (таблицах
выживания).
-
Типы взаимодействия организмов. Теоретически взаимодействие популяций двух видов можно выразить в виде следующих комбинаций символов: 00, --, ++, +0, -0 , +-. Выделяют 9 типов наиболее важных взаимодействий между видами (по Ю. Одуму, 1986):
- Нейтрализм (00)- ассоциация двух видов популяций не сказывается ни на одном из них;
- Взаимное конкурентное подавление (--) - обе популяции взаимно подавляют друг друга;
- Конкуренция из-за ресурсов (--) - каждая популяция неблагоприятно воздействует на другую при недостатке пищевых ресурсов;
- Аменсализм (-0) - одна популяция подавляет другую, но сама при этом не испытывает отрицательного влияния;
- Паразитизм (+ - ) -популяция паразита наносит вред популяции хозяина;
- Хищничество (+ -) - одна популяция неблагоприятно воздействует на другую в результате прямого нападения, но зависит от другой;
- Комменсализм (+0) - одна популяция извлекает пользу от объединения с другой, а другой популяции это объединение безразлично;
- Протокооперация (+ +) -обе популяции получают пользу от объединения;
- Мутуализм (+ +) - связь благоприятна для роста и выживания отдельных популяций, причём в естественных условиях ни одна из них не может существовать без другой.
Примечание: (0) - существенное взаимодействие между популяциями отсутствует; (+) - благоприятное действие на рост, выживание или другие характеристики популяции; ( - ) - ингибирующее действие на рост или другие характеристики популяции.
Тип взаимодействия определённой пары видов может изменяться в зависимости от условий или от последовательных стадий их жизненных циклов.
Девять описанных видов взаимодействий можно свести к двум более обобщенным типам - отрицательным (антибиотическим) и положительным (симбиотическим). В зависимости от состояния экосистемы к ней может быть, применим один из следующих принципов:
- В ходе эволюции и развития экосистемы существует тенденция к уменьшению роли отрицательных взаимодействий за счёт положительных, увеличивающих выживание обоих видов.
- В недавно сформировавшихся или новых ассоциациях вероятность возникновения сильных отрицательных взаимодействий больше, чем в старых.
Взаимодействие популяций может быть взаимно полезным, полезным для одной из них и безразличным для другой. Более того, взаимоотношения могут меняться, они не заданы раз и навсегда.
В случае, когда одна популяция не испытывает влияния другой, т.е. между ними нет взаимодействия, то такая ситуация называется нейтрализмом (белки и лоси в одном лесу). К антибиотическим отношениям можно отнести следующие формы отношений:
- Конкуренцию;
- Паразитизм;
- Хищничество;
- Аменсализм.
К симбиотическим можно отнести следующие формы отношений:
- Собственно симбиоз (протокооперация);
- Мутуализм;
- Комменсализм.
Симбиоз – сожительство, представляет такую форму взаимоотношений, при которой оба партнера или один из них извлекает пользу от другого. Симбиоз - понятие широкое и включает много разных вариантов отношений между видами.
-
Круговорот серы. Сера. Сера является важным составным элементом живого вещества. Большая часть её в живых организмах находится в виде органических соединений. Кроме того, сера входит в состав некоторых биологически активных веществ, а также ряда веществ, выступающих в качестве катализаторов окислительно-восстановительных процессов в организме и активизирующих некоторые ферменты.
Сера представляет собой исключительно активный химический элемент биосферы и мигрирует в разных валентных состояниях в зависимости от окислительно-восстановительных условий среды. Среднее содержание серы в земной коре оценивается в 0,047 %. В природе этот элемент образует свыше 420 минералов.
В изверженных породах сера находится преимущественно в виде сульфидных минералов: пирита FeS2 , пирронита Fe7S8, халькопирита FeCuS2, в осадочных породах содержится в глинах в виде гипсов, в ископаемых углях – в виде примесей серного колчедана и реже в виде сульфатов. Сера в почве находится преимущественно в форме сульфатов; в нефти встречаются её органические соединения.
В связи с окислением сульфидных минералов в процессе выветривания сера в виде сульфат-иона переносится природными водами в Мировой океан, где SO42- занимает второе место по распространению после хлора. Сера поглощается морскими организмами, которые богаче её неорганическими соединениями, чем пресноводные и наземные организмы.
Отметим одну важную особенность малых круговоротов. Применительно к ним понятие круговорота достаточно условно, поскольку природные круговороты не являются замкнутыми. Не всё образуемое при разложении органики неорганическое вещество снова используется живыми организмами. Неиспользуемая его часть образует, в частности, осадочные породы, как в океане, так и на суше, включаясь в большой геологический круговорот. Вещества «не теряются» для биосферы в целом, но уходят из малого круговорота.
-
Круговорот углерода. Углерод в биосфере часто представлен наиболее подвижной формой – углекислым газом. Источником первичной углекислоты биосферы является вулканическая деятельность. Миграция углекислого газа в биосфере Земли протекает двумя путями. Первый путь заключается в поглощении его в процессе фотосинтеза с образованием органических веществ и в последующем захоронении их в литосфере в виде торфа и угля, горных сланцев, рассеянной органики, осадочных горных пород. Так, в далёкие геологические эпохи сотни миллионов лет назад значительная часть фотосинтезируемого органического вещества не использовалась ни консументами, ни редуцентами, а накапливалась и постепенно погребалась под различными минеральными осадками. Находясь в породах миллионы лет, этот детрит под действием высоких температур и давления превращается в нефть, природный газ и уголь, во что именно – зависело от исходного материала, продолжительности и условий пребывания в породах. Теперь мы в огромных количествах добываем это ископаемое топливо для обеспечения потребностей в энергии, а сжигая его, в определенном смысле завершаем круговорот углерода.
По второму пути миграция углерода осуществляется созданием карбонатной системы в различных водоёмах, где СО2 переходит в Н2СО3, НСО31-, СО32-. Затем с помощью растворённого в воде кальция (реже магния) происходит осаждение карбонатов СаСО3 биогенным и абиогенным путями. Возникают мощные толщи известняков. Наряду с этим большим круговоротом углерода существует ещё ряд малых его круговоротов на поверхности суши и в океане.
В пределах суши, где имеется растительность, углекислый газ атмосферы поглощается в процессе фотосинтеза в дневное время. В ночное время часть его выделяется растениями во внешнюю среду. С гибелью растений и животных на поверхности происходит окисление органических веществ с образованием СО2. Особое место в современном круговороте веществ занимает массовое сжигание органических веществ и постепенное возрастание содержания углекислого газа в атмосфере, связанное с ростом промышленного производства и транспорта.
-
Рост популяции. Скорость естественного роста популяции. Кривые роста. Рост популяции - изменение численности ее особей от момента размножения и вселения особей в биоценоз. Определяется как прирост численности в единицу времени. Рост популяции может быть выражен J-образной (экспоненциальной) или S-образной (логистической) кривыми роста. В первом случае кривую отражает уравнение:
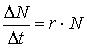 , где
, где
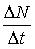 - абсолютная скорость роста численности,
г — биотический потенциал или удельная
скорость роста численности. Решение
этого уравнения — функция
- абсолютная скорость роста численности,
г — биотический потенциал или удельная
скорость роста численности. Решение
этого уравнения — функция
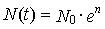 ,
где е —
основание натуральных логарифмов.
График этой функции и есть экспонента.
Если смертность выше рождаемости, то
убывание численности тоже описывается
этим уравнением, но с отрицательным г.
Такой процесс называют экспоненциальным
затуханием
численности.
,
где е —
основание натуральных логарифмов.
График этой функции и есть экспонента.
Если смертность выше рождаемости, то
убывание численности тоже описывается
этим уравнением, но с отрицательным г.
Такой процесс называют экспоненциальным
затуханием
численности.
 С увеличением плотности скорость
роста популяции постепенно снижается
до нуля. При нулевом
росте
популяция стабильна, т. е. размеры ее
не меняются. Отдельные организмы при
этом могут расти и размножаться. Нулевая
скорость
роста означает лишь то, что скорость
размножения, если оно происходит,
уравновешена смертностью. Данная
картина характерна для ряда одноклеточных
и многоклеточных организмов.
Таким
образом, скорость роста популяции в
естественных местообитаниях будет
зависеть от климатических изменений,
от снабжения пищей и от того, ограничено
ли размножение определенным временем
года и др., что должно учитываться при
составлении моделей или их
усовершенствовании.
Второй случай: модель
динамики численности популяции при
ограниченных
ресурсах
предложил в 1845
г. французский математик Ферхюльст.
Уравнение, которое носит его имя,
выглядит так:
С увеличением плотности скорость
роста популяции постепенно снижается
до нуля. При нулевом
росте
популяция стабильна, т. е. размеры ее
не меняются. Отдельные организмы при
этом могут расти и размножаться. Нулевая
скорость
роста означает лишь то, что скорость
размножения, если оно происходит,
уравновешена смертностью. Данная
картина характерна для ряда одноклеточных
и многоклеточных организмов.
Таким
образом, скорость роста популяции в
естественных местообитаниях будет
зависеть от климатических изменений,
от снабжения пищей и от того, ограничено
ли размножение определенным временем
года и др., что должно учитываться при
составлении моделей или их
усовершенствовании.
Второй случай: модель
динамики численности популяции при
ограниченных
ресурсах
предложил в 1845
г. французский математик Ферхюльст.
Уравнение, которое носит его имя,
выглядит так:
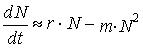 Уравнение Ферхюльста отличается от
уравнения экспоненциального роста
тем, что в правой его части добавляется
выражение mN2.
Это выражение учитывает число встреч
животных, при которых они могут
конкурировать за какой-либо ресурс:
вероятность встречи двух особей
пропорциональна квадрату численности
(точнее, плотности) популяции.
Это же
выражение можно представить в следующем
виде:
Уравнение Ферхюльста отличается от
уравнения экспоненциального роста
тем, что в правой его части добавляется
выражение mN2.
Это выражение учитывает число встреч
животных, при которых они могут
конкурировать за какой-либо ресурс:
вероятность встречи двух особей
пропорциональна квадрату численности
(точнее, плотности) популяции.
Это же
выражение можно представить в следующем
виде:
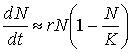 .
Если N
мало по сравнению с k,
то выражение в скобках близко к единице:
при этом уравнение переходит в уравнение
экспоненциального роста. График роста
численности будет при малых N близок к
экспоненте. Когда N
близко к k,
выражение в скобках близко к нулю, т.
е. численность популяции перестает
увеличиваться. Отсюда ясно, что k
в данной модели — это
и есть емкость среды.
При N
больших, чем k,
абсолютный прирост численности
становится отрицательным, и численность
убывает до величины, равной емкости
среды.
.
Если N
мало по сравнению с k,
то выражение в скобках близко к единице:
при этом уравнение переходит в уравнение
экспоненциального роста. График роста
численности будет при малых N близок к
экспоненте. Когда N
близко к k,
выражение в скобках близко к нулю, т.
е. численность популяции перестает
увеличиваться. Отсюда ясно, что k
в данной модели — это
и есть емкость среды.
При N
больших, чем k,
абсолютный прирост численности
становится отрицательным, и численность
убывает до величины, равной емкости
среды. -
-
Биоразнообразие как основа устойчивости экосистем. Биоразнообразие — разнообразие жизни во всех ее проявлениях.
Существует три основных типа биоразнообразия:
- генетическое разнообразие, отражающее внутривидовое разнообразие и обусловленное изменчивостью особей;
- видовое разнообразие, отражающее разнообразие живых организмов (растений, животных, грибов и микроорганизмов). В настоящее время описано около 1,7 млн. видов, хотя их общее число, по некоторым оценкам, составляет до 50 млн.;
- разнообразие экосистем охватывает различия между типами экосистем, разнообразием сред обитания и экологических процессов. Отмечают разнообразие экосистем не только по структурным и функциональным составляющим, но и по масштабу - от микробиогеоценоза до биосферы;
Иногда в отдельную категорию выделяют разнообразие ландшафтов, отражающее особенности территориального устройства и влияние местных, региональных и национальных культур общества.
Экологическая индивидуальность видов растений, сообитающих в одном растительном сообществе в одной экосистеме, позволяет сообществу быстро перестраиваться при изменении внешних условий. Правило Л.Г. Раменского об экологической индивидуальности видов утверждает, что пределы выносливости и зоны оптимумов у разных видов, обитающих совместно, не совпадают. В природе мы находим массу факторов или механизмов, обеспечивающих и поддерживающих высокое видовое разнообразие локальных экосистем. В первую очередь, к таким факторам следует отнести избыточное размножение и перепроизводство семян и плодов. В природе семян и плодов производится в сотни и тысячи раз больше, чем это необходимо, чтобы восполнить естественную убыль в связи с преждевременной гибелью и умиранием от старости. Благодаря приспособлениям к распространению плодов и семян на большие расстояния, зачатки новых растений попадают не только на те участки, которые благоприятны для их произрастания сейчас, но и на такие, условия которых неблагоприятны для роста и развития особей данных видов. Тем не менее, эти семена здесь прорастают, какое-то время существуют в угнетенном состоянии и гибнут. Так происходит до тех пор, пока экологические условия стабильны. Но если условия изменяются, то прежде обреченные на гибель проростки несвойственных этой экосистеме видов начинают здесь расти и развиваться, проходя полный цикл своего онтогенетического (индивидуального) развития. Экологи говорят, что в природе (читай, в биосфере) существует мощное давление разнообразия жизни на все локальные экосистемы. Общий генофонд растительного покрова ландшафтного района – его флора - локальными экосистемами этого района используется наиболее полно именно благодаря давлению биоразнообразия. Таким образом, фактором устойчивости локальной экосистемы является не только разнообразие видов, обитающих в этой локальной экосистеме, но и разнообразие видов в соседних экосистемах, из которых возможен занос диазачатков (семян и спор). Сказанное относится не только к растениям, ведущим прикрепленный образ жизни, но еще в большей степени к животным, могущим перемещаться из одной локальной экосистемы в другую. Следует сказать, что для поддержания равновесия в локальных экосистемах и для их биогеохимической оптимизации важно не таксономическое разнообразие само по себе по принципу «чем больше видов, тем лучше», а разнообразие функциональное, или разнообразие экобиоморф. Мерой функционального разнообразия экосистемы является число экобиоморф и синузий растений, животных, грибов и микроорганизмов. Мерой таксономического разнообразия является число видов, родов, семейств и других высших таксонов. Процессы вымирания видов в биосфере компенсируются процессами видообразования.
-
Демографические характеристики популяции. Популяция – совокупность особей одного вида, занимающих определённую территорию в пределах ареала вида, свободно скрещивающихся друг с другом и в той или иной степени изолированных от жругой популяции. Ценопопуля́ция — совокупность особей вида в пределах одного фитоценоза, занимающего определённое местообитание. Характеристики популяций. Демографические характеристики популяций можно разделить на две группы: статические и динамические. Статические характеристики популяций: Численность - общее число особей в популяции. Может определяться различными способами. Плотность – число особей на единицу площади или объёма, который мы выбираем для учёта. Структура – соотношение различных частей популяции по разным параметрам , выбираемым для учёта. Структура может быть: - демографическая (возрастная) – соотношение разных возрастных групп в популяции;
- половая – соотношение особей разных пород; - пространственная – размещение особей в пространстве. Численность может определяться различными способами. Но зачастую невозможно определить общую численность организмов прямым подсчетом особей. При этом приходится довольствоваться отбором проб и подсчетом количества особей в них. При этом измеряется плотность — количество особей, приходящихся на единицу пространства. Плотность наземных организмов выражают на единицу площади, а популяций планктонных организмов — на единицу объема водной толщи. Иногда плотность оценивают как число встреч на маршруте. Так оценивают птиц по пению в весенний период, млекопитающих по следам зимой.
Выделяют три основных типа пространственных распределений: При случайном распределении нахождение каждой особи никак не зависит от расположения других особей. В природе оно встречается довольно редко.
Регулярное распределение наблюдается в том случае, когда между особями действуют силы отталкивания (например, для них характерна защита индивидуальной территории). Такое размещение часто встречается и в искусственно созданных экосистемах (парках, агросистемах).
При групповом распределении (пятнистом, агрегированном) вероятность нахождения пустых участков и участков с несколькими особями намного выше, чем при случайном распределении. Как и расположение отдельных особей, распределение групп может также быть случайным, регулярным или пятнистым.
Приведенным выше перечнем статические характеристики популяций не исчерпываются. Например, видам, для которых характерна внутрипопуляционная иерархия (порядок подчинения), важна иерархическая структура популяций — отражение статуса отдельных особей и их отношений друг с другом.
Динамические характеристики: Рождаемость – это число новых особей, появляющихся в популяции за единицу времени в расчете на определенное число ее членов. Различают абсолютную (общее число родившихся особей) и удельную (среднее изменение численности на особь за определенный интервал времени). Миграции - регулярное передвижение популяции животных, в ходе которого особи из одной области обитания перемещаются в другую, но затем возвращаются обратно. . Важнейшие динамические характеристики популяции могут быть объединены в следующую простую формулу:
Изменение численности = (рождаемость + иммиграция) — (смертность + эмиграция)
При рассмотрении изменения численности необходимо также учитывать продолжительность жизни особей рассматриваемой популяции. Для организмов каждого вида существует некая максимальная продолжительность жизни особей. Она чаще всего необходима как крайняя точка при построении кривых выживания и демографических таблиц. Так у бактерий она может составлять десятки минут, а у древесных растений — десятки столетий.