

2.5 Біосинтез, шляхи біотрансформації та екскреція холестеролу з організму
Джерелами холестеролу для людини є продукти тварин-ного походження і його біосинтез в організмі. Холестерол синте-зується головним чином у печінці – від 0,5 до 1,0 г за добу (~50% від загальної кількості холестеролу, що синтезується), у кишечнику (~15%) і шкірі. З їжею надходить 0,3-0,5 г холестеролу на добу.
Біологічна роль. Холестерол є незамінним структурним компонентом мембран і зовнішнього шару ліпопротеїнів крові, а також попередником у синтезі кортикостероїдів, статевих гормонів, жовчних кислот, вітаміну D.
2.5.1 Біосинтез холестеролу
Субстратом для синтезу холестеролу є ацетил-КоА. Синтез холестеролу містить 35 послідовних реакцій. Умовно можна виділити 3 основні стадії синтезу: І) перетворення активного ацетату на мевалонову кислоту; ІІ) утворення сквалену з мевалонової кислоти; ІІІ) циклізація сквалену в холестерол. Біосинтез відбувається у мікросомах ЕПР і цитозолі.
І) Перетворення активного ацетату на мевалонову кислоту:
Синтез ацетоацетил-КоА:
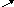 НS-КоА
НS-КоА

 СН3-СО-S-КоА
+ СН3-СО-S-КоА
СН3-СО-СН2-СО-S-КоА
СН3-СО-S-КоА
+ СН3-СО-S-КоА
СН3-СО-СН2-СО-S-КоА
Ацетил-КоА Ацетил-КоА Ацетил-КоА Ацетоацетил-КоА
ацетилтрансфераза
Конденсація ацетоацетил-КоА з ацетил-КоА:
+ Н2О

 СН3-СО-СН2-СО-S-КоА
+ СН3-СО-S-КоА
СН3-СО-СН2-СО-S-КоА
+ СН3-СО-S-КоА
Ацетоацетил-КоА Ацетил-КоА - Н2О
β-окси-β-метилглутарил-К
 оА-
оА-
синтаза (ОМГ-КоА-синтаза)
ОН
│
НООС-СН2-С-СН2-СО-S-КоА + НS-КоА
│
СН3
β-окси-β-метилглутарил-КоА
Синтез мевалонової кислоти:
ОН
│
 НООС-СН2-С-СН2-СО-S-КоА
+ 2НАДФН•Н
НООС-СН2-С-СН2-СО-S-КоА
+ 2НАДФН•Н
│ ОМГ-КоА-редуктаза
СН3
β-окси-β-метилглутарил-КоА
ОН
│
 НООС-СН2-С-СН2-СО-S-КоА
+ НS-КоА
НООС-СН2-С-СН2-СО-S-КоА
+ НS-КоА
│
СН3
Мевалонова кислота
ІІ) Перетворення мевалонової кислоти на сквален:
 Мевалонова кислота
Мевалонова кислота
 АТФ
АТФ
АДФ
5-Фосфомевалонат

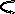 АТФ
АТФ
АДФ
5-Пірофосфомевалонат
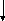
 АТФ
АТФ
АДФ
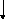
 3-Фосфо-5-Пирофосфомевалонат
3-Фосфо-5-Пирофосфомевалонат
 Фн
Фн
СО2


 Ізопентенілпірофосфат
↔ Диметилалілпірофосфат
Ізопентенілпірофосфат
↔ Диметилалілпірофосфат
(С5) Ізомеризація (С5)
 ФФн
Конденсація
ФФн
Конденсація
Геранілпірофосфат

 (С10)
(С10)

Ізопентенілпірофосфат ФФн
 (С5)
(С5)
Фарнезилпірофосфат
(С15)

 Фарнезилпірофосфат
Відновлювальна
НАДФН+Н+
Фарнезилпірофосфат
Відновлювальна
НАДФН+Н+
(С15) конденсація
НАДФ+, 2ФФн
Сквален
(С30)
ІІІ) циклізація сквалену в холестерол:
НАДФН+Н+ О2 Н2О НАДФН+
С
 квален
Ланостерин
Холестерол
квален
Ланостерин
Холестерол

 (С30)
Скваленоксидоциклаза
(С30)
(С27)
(С30)
Скваленоксидоциклаза
(С30)
(С27)
Вірогідно, проміжні продукти на стадіях перетворення сквалену на холестерол зв'язуються із спеціальним сквален- та стеролтранспортувальним протеїном. Цей білок зв'язує стероли та інші нерозчинні ліпіди, що забезпечує їм можливість участі в реакціях, які протікають у водній фазі клітини. Можливо, що при перетворенні холестеролу на стероїдні гормони та жовчні кислоти, а також при утворенні мембран та ліпопротеїнів, холестерол залишається зв'язаним із холестерол-транспортувальним протеїном.
Реакції біосинтезу холестеролу, що вміщують у себе процеси епоксидації, окисного гідроксилювання, деметилюван-ня, каталізують цитохром-Р450-вмісні монооксигенази, які потребують НАДФН·Н та О2.
Сумарне рівняння біосинтезу холестеролу із Ацетил-КоА:
18 СН3СО-SКоА + 13НАДФН·Н + 3О2 + 18АТФ → С27Н46О + 13НАДФ++ 18 КоАSН + 9СО2 + 18АДФ + 6Н4Р2О7 + 6Н3РО4 + Н2О
Регуляція синтезу холестеролу відбувається на рівні ферменту β-ГОМК-редуктази за рахунок таких молекулярних механізмів:
а) за принципом негативного зворотного зв'язку – холестерол, мевалонова кислота, як кінцеві продукти біосинтезу, зменшують швидкість утворення ензиму. Інгібітором ферменту є також холестеролвмісні ЛПНЩ після їх зв'язування з відповідними рецепторами (рецептори апо-В-100). Споживання холестеролу з їжею гальмує синтез ферменту в печінці, а безхолестеринова дієта активує синтез ензиму;
б) шляхом ковалентної модифікації (фосфорильована форма ферменту – неактивна, дефосфорильована – активна);
в) шляхом ферментної індукції і репресії - залежно від дії біохімічних модуляторів на швидкість синтезу ензиму;
г) гормональна регуляція – інсулін та гормони щитоподібної залози збільшують активність β-ГОМК-редуктази, глюкагон та глюкокортикоїди – зменшують.
Баланс холестеролу в тканинах. Фактори, які впливають на баланс холестеролу в клітині наведені на рис. 17, табл.3.
Клітинна мембрана

Рецептор Регуляція за принципом
Л
 ПНЩнегативного
зворотного зв'язку Синтез
ПНЩнегативного
зворотного зв'язку Синтез
(


 апо-В-100,
Е)_
апо-В-100,
Е)_





 _
_
Л




 ПНЩЕХ
Х
+
ПНЩЕХ
Х
+

АХАТ


Лізосоми Пул вільного
холестеролу ЕХ




Л

 ПНЩ
ЕХ
Х
ПНЩ
ЕХ
Х
Безрецепторний шлях Гідролаза ефірів
холестеролу
Л
 ПНЩХ
ПНЩХ
ЛПДНЩ

 Синтез
стероїдів
Синтез
стероїдів
Х мембрани



 А-1
А-1
ЛХАТ ЕХ
ЛПВЩ
Рисунок 17 – Фактори, які впливають на баланс холестеролу в клітині Позначення: Х- холестерол; ЕХ – ефіри холестеролу; АХАТ – ацил-КоА-холе-стерол ацилтрансфераза; ЛХАТ – лецитин-холестерол ацилтрансфераза;
А-1 – апопротеїно А-1; ЛПНЩ – ліпопротеїни низької щільності; ЛПДНЩ – ліпопротеїни дуже низької щільності
Підвищення вмісту холестеролу в тканинах відбувається за рахунок: 1) захоплення холестеролвмісних ліпопротеїнів
(ЛПНЩ) відповідними рецепторами; 2) захоплення холесте-ролвмісних ліпопротеїнів без участі рецепторів; 3) захоплення вільного холестеролу, який міститься у багатих холестеролом ліпопротеїдах, клітинними мембранами; 4) синтез холестеролу; 5) гідроліз ефірів холестеролу під дією гідролаз ефірів холестеролу (холестерол естераз).
Зменшення вмісту холестеролу спостерігається при:
1) переході холестеролу з мембрани в ліпопротеїни з низьким вмістом холестеролу (ЛПВЩ). Цей перехід регулює фермент ЛХАТ (лецитин-холестерол ацилтрансфераза); 2) естерифікації холестеролу, яке каталізує фермент АХАТ (ацил-КоА-холе-стерол ацилтрансфераза); 3) використанні холестеролу для синтезу інших стероїдів. Інші важливі фактори, які впливають на рівень холестеролу в організмі наведені в таблиці 3.
Таблиця 3. Фактори, які підвищують/знижують рівень холестеролу і
його синтез
|
Підвищують |
Знижують |
|
1 Безхолестеринова дієта підвищує синтез. |
1 Холестерол їжі: за принципом негативного зворотного зв'язку інгі-бує β-ГОМК редуктазу. |
|
2 Споживання великої кількості насичених жирних кислот. |
2 Голодування: інгібування актив-ності β-ГОМК редуктази і підви-щення активності β-ГОМК ліази та синтезу кетонових тіл. |
|
3 Дієта багата на вуглеводи. |
3 Підвищення концентрації жовчних кислот у печінці, які призначаються з лікувальною метою, знижує синтез. |
|
4 Зниження виділення жовчі. |
4 Підвищення абсорбції ліпідів і жовчних кислот із кишечника знижує синтез. |
|
5 Відсутність білків у їжі. |
5 Споживання поліненасичених жирних кислот знижує синтез. |
|
6 Дефіцит перидоксальфосфату. |
6 Циклічний АМФ: інгібує перетворення β-ГОМК редуктази в активну форму. |
|
7 Гормони: інсулін, тиреоїдні гормони, за рахунок активації β-ГОМК редуктази. |
7 Гормони: глюкагон, глюко-кортикоїди знижують синтез. |