

3.Типи бродіння.
ЗАГАЛЬНА ХАРАКТЕРИСТИКА ПРОЦЕСУ БРОДІННЯ
Із трьох принципово можливих способів регенерації АТФ (дихання, бродіння та фотосинтез) бродіння найпростіший.
Бродіння — це такий метаболічний процес, у якому регенерується АТФ, а продукти розщеплення органічного субстрату можуть служити одночасно і донорами, і акцепторами водню. Загальна схема бродіння показана на рис. 6.12. Органічний субстрат є джерелом енергії та вуглецю. Реакції синтезу АТФ є реакціями окиснення. Від окисненого вуглецю клітина позбавляється, виділяючи СО2. Окремі етапи окиснення являють собою дегідрування, за якого водень переноситься на НАД. Акцепторами водню, який міститься у вигляді НАДН, є проміжні продукти розщеплення субстрату. За рахунок НАДН ці проміжні продукти відновлюються, а продукти відновлення виводяться з клітини.

Рис. 6.12. Загальна схема бродіння
У процесі зброджування вуглеводів та інших субстратів утворюються (окремо чи в суміші) такі продукти, як етанол, лактат (молочна кислота), пропіонат, форміат, бутират, сукцінат, ацетат, н-бутанол, 2,3-бутандіол, ацетон, 2-пропанол, СО2 та Н2. Залежно від того, які продукти переважно утворюються, розрізняють спиртове, молочнокисле, маслянокисле, пропіоновокисле, мурашинокисле та оцтовокисле бродіння. Молекулярний кисень у процесах бродіння участі не бере. Більшість мікроорганізмів, які здійснюють бродіння, є облігатними анеробами або факультативними аеробами, здатними рости як у присутності кисню, так і без нього. При цьому кисень пригнічує бродіння, і воно змінюється диханням.
Регенерація АТФ у процесі бродіння
У процесі зброджування глюкози утворюється від одного до чотирьох молей АТФ. У фосфорилюванні на рівні субстрату беруть участь такі три найважливіші реакції:
1,3-Дифосфогліцерат + АДФ -> 3-Фосфогліцерат + АТФ(фосфогліцераткіназа);
Фосфоенолпіруват + АДФ -> Піруват + АТФ (піруваткіназа);
Ацетилфосфат (бутирилфосфат) + АДФ —> Ацетат (бутират) + АТФ (ацетаткіназа).
У більшості мікроорганізмів використовуються перші дві реакції. При цьому необхідні акцептори водню утворюються з пірувату або ацетил-КоА. У процесі зброджування одного моля глюкози утворюється лише два або три моля АТФ, а продуктами бродіння є лактат, етанол, ацетон, бутират, бутанол, 2-пропанол, 2,3-бутандіол, капронат, ацетат, СО2 та Н2,.
При використанні третьої реакції, яка каталізується ацетаткіназою, утворюється додатковий АТФ. Ацетилфосфат утворюється з ацетил-КоА за допомогою фосфотрансацетилази:
Ацетил-КоА + Ф-> Ацетилфосфат + КоА.
Крім того, можливе утворення ацетилфосфату з фосфорильованих сахаридів (ксилулозо-5-фосфату, фруктозо-6-фосфату) за участю фосфокетолази.
Здатність бактерій здійснювати третю реакцію залежить від того, чи можуть вони виділяти молекулярний водень. При перенесенні відновлювальних еквівалентів (електронів) на протони вони можуть виділятися у вигляді молекулярного водню. У цьому разі клітині немає потреби синтезувати акцептори водню. Для того щоб зрозуміти цей механізм, необхідно розглянути механізми вивільнення водню.
Механізми вивільнення водню. Анаеробні бактерії окиснюють піруват до ацетил-КоА двома способами:
Піруват - КоА + 2 Fd -> Ацетил-КоА + FdH + СО2 (Fd — фередоксин)
(фермент піруват:фередоксин-оксидоредуктаза)
Піруват + КоА -> Ацетил-КоА + Форміат
(фермент піруват:форміатліаза).
У першій реакції (є характерною для клостридій) відновлюється фередоксин, окисно-відновний потенціал якого є дуже низьким (-420 мВ), тому за допомогою спеціальної гідрогенази може вивільнюватися водень:
2 FdH -> 2 Fd + Н2 {гідрогеназа).
У другій реакції (є характерною для ентеробактерій) утворюється форміат, окисно-відновний потенціал якого теж є низьким, тому він може розщеплюватися з утворенням Н2:
Форміат —> Н2 + СО2 (гідрогенліазна система).
Враховуючи те, що окисно-відновні потенціали як у FdH, так і у форміату є досить низькими, клітині неважко позбавлятися від відновлювальних еквівалентів, що утворюються при окисненні пірувату до ацетил-КоА.
На відміну від цього, водень, що утворюється у процесі дегідрування гліцеральдегід-3-фосфату у вигляді НАДН, у більшості анаеробних бактерій далі передається на органічні акцептори.
Проте деякі бактерії можуть вивільняти молекулярний водень навіть із НАДН завдяки наявності фермента НАДН: фередоксиноксидоредуктази:
НАДН + 2 Fd -> НАД + 2 FdH.
Як відомо, за допомогою гідрогенази з FdH утворюється водень. Проте ці реакції можуть проходити тільки тоді, коли утворюваний водень безперервно виводиться (споживається). Це зумовлено тим, що такі реакції утворення водню пов'язані з підвищенням потенціалу Н (від -320 мВ для НАДН до -420 мВ для фередоксину), і їх рівновага є несприятливою для виділення водню. Тому організми, які можуть утворювати водень із НАДН, можуть використовувати цей елегантний спосіб позбавлення від водню у формі Н2 тільки тоді, коли існують разом з мікроорганізмами, що безперервно споживають водень. Саме так відбувається у природі. Таке явище називають міжвидовою передачею водню, що є особливою формою симбіозу у мікробних спільнотах.
Звичайно ж, що бактерії, які здатні позбавлятися від зв'язаного з НАД водню, виділяючи його у вигляді Н2, можуть обходитись без реакцій перетворення ацетил-КоА на акцептори для НАДН. Тому вони здатні перетворювати ацетил-КоА на ацетил-фосфат і регенерувати додатковий АТФ шляхом ацетаткіназної реакції. Виділяють вони, головним чином, ацетат і здатні регенерувати до чотирьох молей АТФ.
Роль процесів бродіння у балансі природи
Мікроорганізми, які здійснюють бродіння, відіграють важливу роль у природному кругообігу речовин. Більша частина целюлози, яку споживають рослиножуйні тварини, виводиться з їх організму у неперетравленому вигляді. У разі потрапляння такого целюлозного детриту в анаеробні шари ґрунту або донні осади водойм целюлоза піддається зброджуванню клостридіями і деякими іншими строго анаеробними бактеріями. При цьому утворюються всі названі продукти бродіння, у тому числі майже завжди молекулярний водень. Водень міститься на початку анаеробного харчового ланцюга, головними продуктами якого є метан та/або сірководень. В осадах прісноводних озер та в рубці жуйних тварин водень перетворюється метаноутворювальними бактеріями на метан, а в морських анаеробних екосистемах сульфатовідновлювальні бактерії перетворюють водень і сульфат на сірководень.
СПИРТОВЕ БРОДІННЯ
Основними продуцентами етанолу є дріжджі Saccharomyces cerevisiae та бактерії (Zymomonas mobilis, Leuconostoc mesenteroides, Sarcina ventriculi).
Утворення етанолу дріжджами
Катаболізм глюкози у процесі зброджування її до етанолу та СО2 здійснюється гліколітичним шляхом. Глюкоза окиснюється до пірувату. Перетворення пірувату на етанол проходить у два етапи (рис. 6.13):
1) піруват декарбоксилюється піруватдекарбоксилазою до ацетальдегіду;
2) ацетальдегід відновлюється алкогольдегідрогеназою до етанолу за участю НАДН. При цьому переноситься водень, який утворився під час дегідрування триозофосфату. Окисно-відновний баланс, таким чином, зберігається.
Форми бродінь, відкриті К. Нейбергом. Якщо до дріжджів, які зброджують глюкозу, добавити бісульфіт (він є нетоксич-ним для дріжджів), то з'явиться новий продукт — гліцерин і одночасно знизиться вихід етанолу та СО2. Ацетальдегід зв'язується бісульфітом з утворенням ацетальдегідсульфіту і тому не може служити акцептором водню — етанол не утворюється. Замість ацетальдегіду таким акцептором водню є діоксиацетонфосфат. Він відновлюється до гліцерин-3-фосфату і дефосфорилюється до гліцерину. Бродіння у присутності бісульфіту використовують у промисловості для одержання гліцерину. Це друга форма бродіння за Нейбергом.
Якщо у процесі бродіння у розчин добавити бікарбонат натрію або двозаміщений фосфорнокислий натрій, також утворюється гліцерин, оскільки ацетальдегід перетворюється в результаті реакції дисмутації на етанол та оцтову кислоту і тому не може служити акцептором водню. Це третя форма бродіння за Нейбергом. Пертою формою бродіння є нормальне дріжджове бродіння.
Ефект Пастера. Зброджування дріжджами глюкози — процес анаеробний, хоча дріжджі є аеробними мікроорганізмами. В анаеробних умовах бродіння відбувається дуже інтенсивно, але відзначається слабкий ріст. Під час аерації бродіння послаблюється, починається дихання і активний ріст. У деяких дріжджів бродіння можна майже пригнітити посиленою аерацією (ефект Пастера). Л. Пастер відкрив цей ефект понад сто років тому. Він властивий не тільки дріжджам, а й усім факультативно аеробним клітинам, у тому числі й клітинам тканин вищих тварин. Утворення етанолу бактеріями
Відомий для дріжджів шлях утворення етанолу (гліколіз, піруватдекарбоксилазна реакція, відновлення ацетальдегіду) з усіх досліджених бактерій виявлений тільки у Sarcina uentriculi.
Бактерія Zymomonas mobills розщеплює глюкозу КДФГ-шляхом, піруват розкладається у піруватдекарбоксилазній реакції на ацетальдегід і СО2, ацетальдегід відновлюється до етанолу (див. рис.6.13). Єдиними продуктами бродіння у цьому разі є етанол, вуглекислий газ і невеликі кількості молочної кислоти. У процесі бродінь, що здійснюються деякими видами ентеро-бактерій і клостридіями, етанол є побічним продуктом. Попередник етанолу — ацетальдегід — утворюється у цьому разі не з піру-вату, а шляхом відновлення ацетил-КоА.
Зовсім іншим шляхом утворюють етанол гетероферментативні молочнокислі бактерії Leuconostoc mesenteroides (див. рис. 6.13). Глюкоза розкладається пентозофосфатним шляхом до пентозо-фосфату. Ксилулозо-5-фосфат за участю фосфокетполази перетворюється на ацетилфосфат і гліцеральдегід-3-фосфат:
Ксилулозо-5-фосфат + Ф —> Ацетилфосфат + Гліцеральдегід-3-фосфат.

Рис.6.13. Спиртове бродіння
Ацетилфосфат відновлюється ацетальдегіддегідрогеназою та алкогольдегідрогеназою до етанолу. Інший продукт розщеплення глюкози — гліцеральдегід-3-фосфат — відновлюється до лактату через піруват.
МОЛОЧНОКИСЛЕ БРОДІННЯ І РОДИНА LACTOBACILLACEAE
Характеристика молочнокислих бактерій
Молочнокислі бактерії об'єднують у родину LactobacilІасеае. Ця група є морфологічно гетерогенною, але має спільні фізіологічні ознаки. Всі бактерії є грампозитивними, не утворюють спор (за винятком Sporolactobacillus inulinus), переважно нерухливі. Всі вони використовують вуглеводи як джерело вуглецю та енергії і виділяють молочну кислоту. На відміну від ентеробактерій, які теж виділяють лактат, молочнокислі бактерії здатні тільки до бродіння, вони не містять гемопротеїнів (цитохромів і каталази). Проте представники родини Lactobacillaceae можуть рости у присутності кисню, вони все-таки є аеротолерантними.
Ще однією особливістю молочнокислих бактерій є потреба в ростових факторах. Для росту більшості цих бактерій потрібен ряд вітамінів (тіамін, пантотенова, нікотинова, фолієва кислоти, біотин), амінокислот, а також пуринів і піримідинів. Вирощують молочнокислі бактерії переважно на складних середовищах, що містять достатньо високі концентрації дріжджового автолізату, томатного соку, молочної сироватки і навіть крові. Отже, молочнокислі бактерії є своєрідними "метаболічними інвалідами", які, очевидно, в силу своєї спеціалізації (ріст на молоці та інших багатих середовищах) втратили здатність до синтезу багатьох метаболітів. З іншого боку, їм притаманна здатність, якої немає в інших мікроорганізмів: вони можуть використовувати молочний цукор (лактозу). Лактоза зустрічається тільки у ссавців, у рослинному царстві її немає. Лактоза являє собою дисахарид, який перш ніж катаболізуватися, повинен бути розщеплений на глюкозу та галактозу (фермент в-галактозидаза). У зв'язку з тим, що молочнокислі бактерії виділяють велику кількість молочної кислоти, середовище для їх культивування повинно бути забуференим.
Молочнокислі бактерії ніколи не зустрічаються в ґрунті або водоймах. Місця їх існування такі: молоко й місця його переробки, молочні продукти (Lactobacillus lactis, L. bulgaricus, L. casei, L. fermentum, L. brevis, Streptococcus lactis);рослини й рослинні залишки (Lactobacillus plantarum, L. delbruekii, L. fermentum, L. brevis, Streptococcus lactis, Leuconostoc mesenteroides);кишечник і слизові оболонки людини й тварин (Lactobacillus acidophilus, Bifidobacterium bifidum, Streptococcus faecalis, S. bouts, S. pneumoniae).
Серед стрептококів зустрічаються паразити крові — досить вірулентні збудники захворювань.
Залежно від того, які продукти утворюються у процесі зброджування глюкози (тільки молочна кислота або також і інші органічні продукти та СО2), молочнокислі бактерії поділяються на гомо- та гетероферментативні.
Гомоферментативне молочнокисле бродіння
Гомоферментативні молочнокислі бактерії утворюють тільки одну молочну кислоту (вона становить не менше 90 % усіх продуктів бродіння). Глюкоза катаболізується гліколітичним шляхом. Водень, який відщеплюється під час дегідрування гліцеральдегід-3-фосфату у вигляді НАДН, передається на піруват рис. 6.14). У присутності лактатдегідрогенази піруват відновлюється до лактату. Лише невелика частина пірувату декарбоксилюється та перетворюється на оцтову кислоту, етанол і СО2, а також адетоїн. Гомоферментативне молочнокисле бродіння здійснюють стрептококи, серед лактобацил — Lactobacillus lactis, L. bulgaricus, L. delbruckii, L. acidophilus.
Гетероферментативне молочнокисле бродіння
У гетероферментативних молочнокислих бактерій (на відміну від гомоферментативних) відсутні такі ферменти гліколізу, як альдолаза та
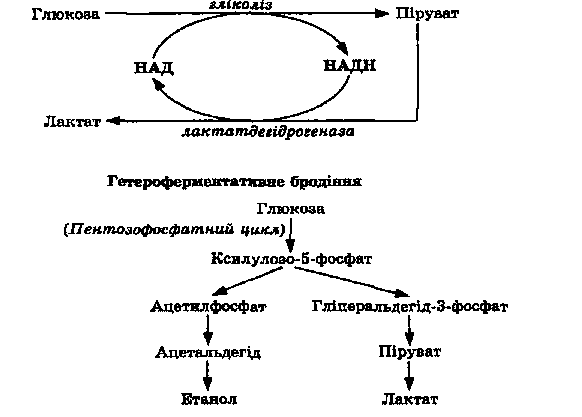
Рис. 6.14. Молочнокисле бродіння
триозофосфатізомераза. Початкове розщеплення глюкози у них відбувається лише пентозофосфатним шляхом (див. рис6.14). Рибулозо-5-фосфат під дією епімерази перетворюється на ксилулозо-5-фосфат, який в результаті реакції, що каталізується пентозофосфокетолазою, розщеплюється з утворенням глідеральдегідфосфатутаацетилфосфату. З гліцеральдегідфосфату через піруват утворюється молочна кислота, а ацетилфосфат відновлюється через ацетил-КоА та ацетальдегід до етанолу. Такий тип бродіння є характерним для Leuconostoc mesenteroides.
Інші гетероферментативні бактерії переводять частково чи повністю ацетилфосфат в оцтову кислоту, що супроводжується утворенням АТФ. У цьому разі надлишок водню передається на глюкозу, яка відновлюється до маніту. З гліцеральдегідфосфату через піруват утворюється лактат.
Зброджування фруктози здійснюється за рівнянням
3 Фруктоза -» Лактат + Ацетат + СО2 + Маніт.
Фруктоза при цьому служить акцептором надлишкових відновлювальних еквівалентів і відновлюється до маніту.
Гетероферментативне молочнокисле бродіння здійснюють Leuconostoc mesenteroides, L. fermentum, L. brevls, Bifidobacterium bifidum.
Бродіння, яке здійснюється Bifidobacterium bifidum. Ця бактерія отримала назву за свою V- та Y-подібну форму (лат. bif idus — роздвоєний). Вона відома тим, що переважає в кишечнику немовлят. Всі представники роду Bifidobacterium є строгими анаеробами, вони ростуть в атмосфері, яка містить 10 % СО2. Біфідобактерії розщеплюють глюкозу на молочну кислоту та ацетат. Причому вони не мають ні альдолази, ні глкжозо-6-фосфатдегідрогенази. У цих бактерій глюкоза перетворюється на фруктозо-6-фосфат гліколітичним шляхом, а потім фруктозо-6-фосфат за допомогою активних фосфокетолаз перетворюється на ацетилфосфат і ксилу-лозо-5-фосфат. Ксилулозо-5-фосфат у свою чергу дає ацетилфосфат і гліцеральдегідфосфат. Ацетилфосфат відновлюється до ацетату, а гліцеральдегідфосфат через піруват — до лактату.
Використання молочнокислих бактерій
Розмножуючись, молочнокислі бактерії сильно знижують рН (до значень, менших за 5,0) і тим самим пригнічують ріст інших анаеробних бактерій. Завдяки такій стерилізувальній і консервувальній дії вони використовуються в домашньому та сільському господарстві, молочній промисловості. У домашньому господарстві, насамперед для квашення капусти, у молочній промисловості — для виготовлення заквасок з метою одержання молочнокислих продуктів (сметана, кефір, йогурти, ряжанка та ін).
Виготовлення силосу. Для виготовлення силосу використовуються молочнокислі бактерії, які живуть на рослинах та їх залишках. Для цього використовують листя цукрових буряків, кукурудзи, картоплі, трави та люцерни. Рослинну масу пресують, добавляють мелясу (з метою підвищення співвідношення вуглець: азот), підкислюють з метою створення сприятливих для росту молочнокислих бактерій умов. У таких умовах відбувається контрольоване молочнокисле бродіння.
ПРОПІОНОВОКИСЛЕ БРОДІННЯ ТА ПРОПІОНОВОКИСЛІ БАКТЕРІЇ
Характеристика пропіоновокислих бактерій
Ці бактерії існують у рубці та кишечнику жуйних тварин (корови, вівці), де вони беруть участь в утворенні жирних кислот, головним чином, пропіонової та оцтової. Завдяки цим бактеріям молочна кислота, яка утворюється в результаті різних видів бродінь, перетворюється на пропіонову. Пропіоновокислі бактерії не зустрічаються в молоці, у ґрунті, у водоймах. Рід Propionibacterium складається з грампозитивних нерухомих палички, які не утворюють спор. Ці бактерії не переносять присутності кисню, ростуть в анаеробних умовах, регенерують АТФ за рахунок бродіння. На основі таких ознак їх довго вважали організмами, які облігатно здійснюють бродіння. Проте у них були виявлені такі ферменти, як каталаза та цитохроми. Пізніше було встановлено, що представники роду Propionibacterium здатні рости і в аеробних умовах (за слабкої аерації), але при цьому треба мати на увазі, що кисень все-таки є для них токсичним. Таким чином, щодо кисню пропіоновокислі бактерії є мікроаеротолерантними організмами. Крім роду Propionibacterium, до пропіоновокислих бактерій відносять Veillonella alcalescens, Clostridium propionicum, представників родів Selenomonas, Micromonospora. В анаеробних умовах ці бактерії зброджують Сахарозу, глюкозу, лактозу, пентози, а також малат, лактат, гліцерин та інші субстрати з утворенням пропіонової кислоти. Розщеплення гексоз здійснюється гліколітичним шляхом.
Утворення пропіонової кислоти
Відновлення лактату чи пірувату до пропіонової кислоти відбувається метилмалоніл-КоА-шляхом (рис.6.15) Спочатку піруват перетворюється на оксалоацетат, який відновлюється через малат і фумарат до сукцинату. Транспорт електронів на дій ділянці спряжений з фосфорилюванням (фумаратне дихання). Далі сукцинат трансформується у сукциніл-КоА та метилмалоніл-КоА, у процесі декарбоксилювання якого утворюється пропіоніл-КоА.
Таким шляхом пропіонат утворюється у більшості пропіоновокислих бактерій, а також у Velllonella alcalescens, Selenomonas ruminantium Пропіоніл-КоА може утворюватись також акриолоїл-КоА-шляхом(див рис.6.15). Проміжним продуктом при цьомує похідне акрилової кислоти — акрилоїл-КоА. Так відбувається утворення пропіонату у Clostridlum proplonicum, Bacteroides ruminicola, Megasphaera elsdenii.
.

Рис. 6.15. Пропіововокисле бродіння
МУРАШИНОКИСЛЕ БРОДІННЯ ТА РОДИНА ENTEROBACTERlACtAE
Характеристика ентеробактерій
Деякі мікроорганізми, які утворюють у процесі бродіння кислоти, об'єднують в одну фізіологічну групу на тій підставі, що характерним, хоча і не основним, продуктом бродіння у них є мурашина кислота. Оскільки деякі типові представники цієї групи існують у кишечнику, вся родина має назву Enterobacteriaceae. Це грамнегативні, активно рухливі, що не утворюють спор палички з перитрихіальним джгутикуванням. Факультативні аероби, здатні одержувати енергію як у процесі дихання (аеробні умови), так і у процесі бродіння (анаеробні умови). Дуже невибагливі щодо живлення: ростуть на простих мінеральних середовищах.
Escherichia coli — мешканець кишечника, проте за кількістю не є основним (у кишковій мікрофлорі переважають Bacteroides і Bifidobacterium). Ця бактерія тривалий час зберігає життєздатність і поза кишечником. Цим користуються для виявлення забрудненості питної води фекаліями.
До нормальної кишкової мікрофлори належать також Proteus vulgarls, Enterobacter aerogenes. До родини Enterobacteriaceae належать бактерії роду Erwinla — фітопатогенні бактерії, які уражують стебла, листя та корені рослин.
Особливості бродіння Escherichia coli. Такими особливостями є:
1) розщеплення пірувату на ацетил-КоА та форміат (мурашина кислота); 2) розкладання форміату на СО2 та Н2; 3) відновлення ацетил-КоА до етанолу; 4) утворення сукцинату як продукту фумаратного дихання; 5) відсутність здатності утворювати з пірувату ацетоїн і бутандіол (рис. 6.16).
Особливості бродіння Enterobacter aerogenes. Ацетоїн утворюється з двох молекул пірувату (див. рис.6.16). Цей про цес складається з двох реакцій декарбоксилювання з виділенням CO2. Саме таке інтенсивне газовиділення дало назву цій бактерії. Проміжним продуктом є ацетиллактат. 2,3-бутандіол утворюється при відновленні ацетоїну за участю НАДН.
МАСЛЯНОКИСЛЕ ТА АЦЕТОНО-БУТИЛОВЕ БРОДІННЯ. КЛОСТРИДІЇ
Масляна кислота (бутират), н-бутанол, ацетон, пропанол є типовими продуктами бродіння анаеробних спороутворювальних бактерій (клостридій). Разом з клостридіями розглянемо також деякі спеціалізовані види бактерій, які утворюють у процесі зброджування аналогічні продукти.
Характеристика бактерій роду Clostridium
Цей рід належить до родини Васillасеае. Як і інші представники цієї родини (Bacillus, Sporolactobacillus, Sporosarcina), клостридії є грампозитивними. Рухливі (перитрихально розміщені джгутики), вегетативні

Рис. 6.16. Мурашинокисле бродіння
клітини паличкоподібні, спори змінюють форму материнської клітини, оскільки їх діаметр є більшим за товщину самої клітини. Спори є терморезистентними.
Клостридії відзначаються різко вираженим бродильним типом метаболізму. Вони ростуть тільки в анаеробних умовах, хоча серед них є аеротолерантні види. Із запасних речовин характерними є крохмалеподібні полісахариди. Температурний оптимум становить 30-40 °С, є термофільні види (Clostridium thermoaceticum). Оптимальним рН є нейтральне або лужне.
За здатністю використовувати різні субстрати клостридії можна поділити на дві групи: цукролітичні та пептолітичні.
Зброджування глюкози клостридіями
Катаболізм глюкози здійснюється гліколітичним шляхом. Водень, що утворюється у процесі дегідрування гліцеральдегідфосфату, переноситься на органічні кислоти або кетони, які утворюються з пірувату або ацетил-КоА. Типовим для клостри-дій процесом зброджування глюкози є бродіння, характерне для Clostridium butyricum ї Clostridium acetobutyricum (рис. 6.17).

Рис. 6.17. Зброджування глюкози Clostridium butyricum та Clostridium acetobutyricum
Утворення бутирату починається з конденсації двох молекул ацетил-КоА і утворенням ацетоацетил-КоА, який потім відновлюється до в-оксибутирил-КоА і бутирил-КоА. із ацетил-КоА за участю фосфотрансацетилази та ацетаткінази може утворюватись вільний ацетат, а також молекула АТФ. У міру утворення ацетату та бутирату і зумовленого накопиченням цих кислот підкислення середовища починають синтезуватись ферменти, що каталізують утворення ацетону та бутанолу. Ацето-ацетил-КоА трансформується в ацетоацетат, декарбоксилюван-ня якого дає ацетон. За відновлення ацетону утворюється про-панол. Відновлення бутирил-КоА дає бутанол. У процесі зброджування глюкози клостридіями може також утворюватись і етанол (за відновлення ацетил-КоА). Молекулярний водень може виділятись із НАДН, одержаного дегідруванням гліцеральдегідфосфату та окисненням пірувату. Чим більше молекулярного водню буде одержано, тим менше потрібно синтезувати органічних акцепторів водню (ацетоацетил-КоА).
Оскільки ацетон, 2-пропанол та бутанол є важливими органічними розчинниками, бродіння, яке здійснюється клостридіями, має велике технічне значення.
Зброджування клостридіями субстратів, відмінних від глюкози
Етанол та ацетат. Clostridium kluyveri здатна зброджувати суміш етанолу й ацетату до бутирату та молекулярного водню (рис. 6.18). Ацетат є додатковим акцептором водню, він утворюється у процесі бродіння. АТФ синтезується тільки в ацетаткіназній реакції.

Рис. 6.18.. Зброджування етанолу та ацетату Clostridium kluyveri
Глутамінова кислота. Зброджування глутамінової кислоти здійснюють бактерії Clostridium tetanomorphum. Це бродіння привернуло до себе увагу тим, що під час його дослідження була встановлена біохімічна функція вітаміну В12. У процесі зброджування глутамату утворюються бутират, ацетат, аміак, СО2 та молекулярний водень {рис. 6.19). Шлях катаболізму глутамату складається з ряду незвичних реакцій — утворення метиласпарагінової кислоти (за участю кофермента — похідного вітаміну В12). На наступній стадії — утворення метилфумарату — відбувається дезамінування. Метилмалат розщеплюється на ацетат і піруват. Ацетат виділяється, а піруват перетворюється на СО2 та бутират, як показано на рис. 6.19.

Рис.6.19.. Зброджування глутамату Clostridium tetan.omorphum
Спряжене зброджування двох амінокислот (реакція Стікленда). Пептолітичні клостридії гідролізують білки і потім використовують амінокислоти. Більшість амінокислот зброджуються не окремо, а разом з якими-небудь іншими. Як встановив американський учений Л.Г. Стік ленд (1934 p.), Clostridium sporogenes зброджує суміш аланіну та гліцину, але не може використовувати жодну з цих кислот окремо. Продуктами бродіння є ацетат, аміак і СО2. Аланін є донором водню, а гліцин — акцептором. Амінокислота-донор дезамінується і перетворюється на оксокислоту. Оксокислота в результаті окиснювального декарбоксилювання перетворюється на жирну кислоту. Цей етап спряжений з фосфорилюванням і являє собою реакцію, яка постачає енергію.
Клостридії — збудники захворювань
Деякі пептолітичні клостридії можуть бути збудниками інфекційних хвороб у разі поранень (газова гангрена та правець), а також причиною харчових отруєнь. Якщо спори Clostridium histolyticum і С. septicum (спори цих бактерій дуже поширені в ґрунті) потрапляють у відкриту рану, до якої немає доступу повітря або в якій аеробні бактерії, розмножуючись, створюють анаеробні умови, то клостридії починають рости, розщеплювати за допомогою протеаз коллаген та інші білки і здійснювати бродіння.
Найважчу форму харчового отруєння — ботулізм — спричиняє Clostridium botulinum. Ця ґрунтова бактерія може розвиватися в недостатньо простерилізованих м'ясних продуктах і бобових консервах. Утворюваний нею токсин у разі споживання ураженого продукту може спричиняти смерть внаслідок нервового паралічу, зокрема паралічу дихання. Проте цей токсин є термолабільним та інактивується при кип'ятінні впродовж 15 хвилин.
ГОМОАЦЕТАТНЕ БРОДІННЯ. СО2 ЯК АКЦЕПТОР ВОДНЮ
Деякі клостридії (С. thermoaceticum, C. formicoaceticum, С. cyllndrosporum) здатні переносити водень, відщеплений від субстрату в початкових реакціях окиснення, тільки на Со2, утворюючи при цьому ацетат:
8[Н] + 2СО2 -» СН3-СООН + 2Н2О.
Глюкоза окиснюється гліколітичним шляхом до пірувату. Піруват за допомогою піруват: фередоксин-оксидоредуктази розщеплюється на ацетил-КоА, СО2 та відновлений фередоксин. Із ацетил-КоА через ацетилфосфат утворюється ацетат і АТФ. Акцепторами водню є СО2, при цьому також утворюється ацетат.
ЗБРОДЖУВАНІ ТА НЕЗБРОДЖУВАНІ ПРИРОДНІ СПОЛУКИ
Більшість природних сполук, що складаються з вуглецю, водню, кисню та/або азоту, піддаються зброджуванню в анаеробних умовах. Зброджуються полісахариди, гексози, пентози, багатоатомні спирти, органічні кислоти, амінокислоти (за винятком ароматичних), пурини та піримідини.
Не можуть зброджуватися насичені аліфатичні та ароматичні вуглеводні, стероїди, каротиноїди. Однією з причин цього є те, що ці сполуки можуть окиснюватися тільки в присутності молекулярного кисню за участю оксигеназ. Очевидно, завдяки такій стабільності вуглеводні так довго зберігаються на нафтових родовищах.
Узагальнена схема найважливіших бродінь показана на рис.6.20

.Рис. 6.20. Узагальнена схема найважливіших типів бродінь
НЕПОВНІ ОКИСНЕННЯ
Більшість аеробних мікроорганізмів у процесі дихання окиснюють органічні поживні речовини до СО2 та води. Оскільки в молекулі СО2 досягається найвищий ступінь окиснення вуглецю, то відбувається повне окиснення. Цей тип дихання відрізняють від неповних окиснень, за яких як продукти обміну виділяються частково окиснені органічні сполуки.
Кінцевими продуктами неповних окиснень можуть бути оцтова, глюконова, фумарова, лимонна, молочна кислота та інші сполуки. Оскільки ці продукти подібні до тих, що утворюються у процесі бродіння, неповні окиснення називають "окиснювальним бродінням".
Неповними окисненнями з біотехнології є такі важливі процеси, як утворення органічних кислот бактеріями та грибами; утворення амінокислот бактеріями; трансформація речовин мікроорганізмами; утворення вторинних метаболітів (антибіотиків, полісахаридів, вітамінів, лектинів та ін.).
Утворення оцтової кислоти та оцтовокислі бактерії
Усім оцтовокислим бактеріям притаманна здатність утворювати кислоти неповним окисненням цукрів або спиртів. Оцтовокислі бактерії — це грамнегативні палички з перитрихально (Acetobacter) чи полярно (Gluconobacter) розміщеними джгутиками, які здатні до деякої рухливості. Характеризуються високою стійкістю до кислот. У природних умовах вони, як правило, існують на рослинах. Серед оцтовокислих бактерій розрізняють групи peroxydans і suboxydans. Група peroxydans — організми, які накопичують кислоту тільки як проміжний продукт. Типовим представником є Gluconobacter oxydans. У організмів групи suboxydans оцтова кислота не піддається подальшому окисненню (Acetobacter aceti, Acetobacter pasterianum). Між цими двома групами існує багато проміжних форм, які здатні окиснювати оцтову кислоту дуже повільно. Більшість оцтовокислих бактерій потребує для росту складних поживних середовищ.
Оцтовокислі бактерії здатні рости на середовищі з етанолом. Етанол окиснюється НАД+-залежними алкоголь- та ацетальде-гіддегідрогеназами з утворенням ацетату та двох молекул НАДН. Утворений НАДН використовується для одержання АТФ у дихальному ланцюгу. Крім етанолу, оцтовокислі бактерії окиснюють велику кількість інших спиртів до відповідних кислот і кетонів:
Пропанол —> Пропіонат.
Ізопропанол —> Ацетон.
Гліцерин — >Діоксиацетон.
Глюкоза -> Глюконат.
Глюконат —> 5-Оксоглюконат.
Ці організми не здатні до синтезу катаболічних ферментів, необхідних для високоактивного розщеплення цих субстратів. У результаті кислоти й спирти виділяються у середовище. Оцтовокислі бактерії характеризуються наявністю ПХХ-залежної глюкозодегідрогенази.
Особливий інтерес викликає здатність оцтовокислих бактерій окиснювати D-сорбітол до L-сорбози. Сорбоза потрібна у великих кількостях для синтезу вітаміну С.