

3.2. Спороутворення (споруляція)
Спори утворюються всередині бактеріальної клітини. Цей процес починається з накопичення білкового матеріалу, тому показник заломлення у місці утворення спори збільшується. При цьому відбувається споживання запасних речовин (полі-в-гідроксимасляної кислоти у аеробів і полісахаридів у анаеробів). Упродовж перших 5 год спороутворення значна частина білків материнської клітини розкладається. При цьому утворюється специфічна для спор речовина — дипіколінова кислота, яка не зустрічається у вегетативних клітинах. У ході синтезу дипіколінової кислоти відбувається поглинання іонів кальцію; очевидно, в зрілих спорах ця кислота міститься у вигляді хелату з кальцієм і може становити 10—15 % сухої речовини спор. Дипіколінова кислота міститься тільки в терморезистентних спорах.
Спороутворення є одним з найскладніших процесів диференціації бактеріальної клітини. Воно починається з особливого нерівномірного поділу клітини (рис.2.11).
У результаті інвагінації цитоплазматичної мембрани частина протопласта відділяється від материнської клітини. Цей протопласт містить частину ядерного матеріалу — один геном. Утворення клітинної стінки між обома протопластами (як за звичайного поділу клітини) не відбувається. Замість цього протопласт чохлика. Екзоспорій міститься тільки у не багатьох бактерій (наприклад у Bacillus cereus). майбутньої спори оточується, мовби обростає, плазматичною мембраною материнської клітини. В результаті навкруги протопласта розміщуються дві плазматичні мембрани і кожна з них бере участь у синтезі стінки спори. Мембрана протопласта спори синтезує назовні від себе стінку зародкової клітини, а мембрана, яка походить від материнської клітини, синтезує всередину кору спори (кортекс). Кортекс складається з багатошарового пептидогліканового остова, який відрізняється від каркаса стінок вегетативної клітини ступенем зшивання. Зовнішню оболонку спори утворює материнська клітина. Ця оболонка складається з поліпептидів. Материнська клітина утворює також ще один додатковий поліпептидний шар — екзоспорій, який оточує спору у вигляді особливого чохлика.Єкзоспорій містеться у не багатьох бактерій.

Рис. 2.11. Спороутворення і будова зрілої спори:
А, Б — процес відділення протопласта спори; В, Г,Д — утворення передспори; Е — зріла спора; 1 — цитоплазма; 2 — плазматична мембрана; 3 — клітинна стінка зародкової клітини; 4 — кора спори; 5,6 — відповідно внутрішня і зовнішня оболонки спори; 7 — екзоспоріум
Індукція спороутворення. Спори не є обов'язковою стадією життєвого циклу бактерій. За сприятливих умов бактерії можуть необмежений час розмножуватися поділом, як вегетативні клітини. Утворення спор починається лише тоді, коли не вистачає поживних речовин або коли в надлишку накопичуються продукти обміну. Інакше кажучи, воно відбувається лише тоді, коли для цього створюються певні умови. Висихання не стимулює спороутворення. Якщо помістити вегетативні клітини в дистильовану воду, можна спостерігати "ендотрофну споруляцію", тобто утворення спор за рахунок внутрішньоклітинних запасних речовин. У таких випадках утворення спор, очевидно, викликане браком екзогенного субстрату. Індукція спороутворення відбувається впродовж декількох годин. Якщо, наприклад, до суспензії вегетативних клітин Bacillus cereus у перші 5 год після внесення їх у дистильовану воду добавити глюкозу, то утворення спор припиниться: добавлення субстрату пригнічує споруляцію. Але якщо глюкозу добавити пізніше, ніж через 6 год, пригнічення спороутворення не відбувається. Індукція спороутворення продовжується, і вже через 10-13 год після внесення у воду близько 90 % клітин утворюють спори. Отже, споруляція регулюється зовнішніми факторами.Кількість клітин, що утворюють спори, у багатьох випадках збільшується після добавлення в середовище солей марганцю.
Здатність утворювати ендоспори поступово втрачається за багаторазових пересівів вегетативних клітин. Оскільки суспензія спороутворювальних бактерій містить і спори, і вегета-
тивні клітини, перед кожним пересіванням культуру, як правило, піддають короткочасному кип'ятінню. Це сприяє зберіганню чи підвищеній здатності клітин утворювати спори.
Властивості зрілих спор. Спори вивільнюються при автолізі материнських клітин. Зрілі спори не виявляють ніякої метаболічної активності. Вони надзвичайно стійкі до дії високих температур, різного роду опромінень, хімічних агентів. Терморезистентність зумовлена низьким вмістом води у спорах — 15 % (стільки само, скільки в сухому казеїні або у вовні) і приблизно пропорційна вмісту в них дипіколінової кислоти.
Проростання спор. У відповідних сприятливих умовах більшість спор проростають. Відповідне попереднє оброблення, певні умови зберігання та прогрівання можуть підвищити "схожість" спор — збільшити процент проростання. У випадку Bacillus subtills оптимальними умовами для стимуляції проростання спор вважають семиденний період спокою і п'ятихвилинне прогрівання у воді при 60 °С. Інші спори можуть бути активовані короткочасним кип'ятінням (10 хв, 100 °С). Теплове оброблення спор повинно проводитися безпосередньо перед висіванням, оскільки процес активації є зворотним.
Проростанню спор передує поглинання ними води та набухання. Для проростання активованих спор у ряді випадків необхідною є наявність глюкози, амінокислот, нуклеозидів та інших речовин. У ході проростання відбуваються глибокі фізіологічні зміни: дихання та ферментативна активність швидко підвищуються, починається виділення амінокислот, дипіколінової кислоти та пептидів. У процесі проростання втрати сухої речовини досягають 25-30 %. Під час проростання спори втрачають термостійкість. Ростова трубка, яка виходить із спори, оточена дуже тонкою, не до кінця сформованою клітинною стінкою, так що в протопласт може проникати навіть ДНК. Ростова трубка може утворюватись як у полярному, так і латеральному положенні {рис.2.12). В одному випадку оболонка спори при цьому розривається, в іншому — ростова трубка її проколює.
Тривалість життя спор. Бактерії у вигляді спор можуть тривалий час перебувати у стані анабіозу. Так, у зразках ґрунту, що зберігалися 50-100 років, були виявлені спори бацил. За даними такого роду експериментів, у сухому ґрунті за 50 років зберігання до 90 % спор втрачають життєздатність. Але при цьому 1 т сухого ґрунту навіть через 1000 років міститиме життєздатні спори.
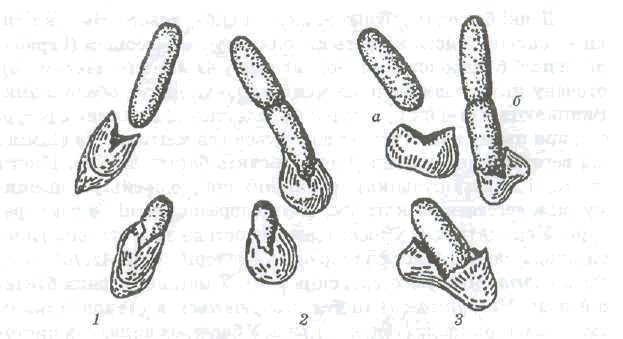
Рис. 2.12. Проростання спори:
1,2 — полярне; 3 — латеральне; а — Bacillus cereus; б — Bacillus subtilis
У сухому стані багато бактерій (якщо не всі з них) упродовж багатьох років зберігають життєздатність. Для консервації бактерій у колекціях культур вегетативні клітини, як правило, піддають ліофільному висушуванню (висушуванню з замороженого стану) і зберігають при кімнатній температурі або низьких температурах у вакуумі. Бактерії, які не витримують ліофілі-зації, зберігаються упродовж багатьох років у вигляді суспензії при температурі рідкого азоту.