

Глава 5. Методы выявления и оценки генотоксикантов
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Таблица 5.2.4 |
||||
|
|
Сравнение данных по выживаемости |
|
|
|
|
||||||||
|
|
и частоте мутаций у хлореллы |
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
Количество |
Число сте- |
Выживаемость, |
|
Частота мутаций, |
||||||||
|
Варианты |
пеней сво- |
|
|
W, % |
|
|
|
M ,% |
|
||||
|
чашек, n |
|
|
|
|
|
|
|||||||
|
|
боды, ν |
X |
|
m |
|
X |
|
m |
|
||||
|
|
|
|
|
|
|
||||||||
|
Контроль |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Опыт |
|
|
|
|
|
|
|
|
|
|
|
|
|
5.3. Определение генотоксического действия факторов окружающей среды с использованием Allium cepa
в качестве тест-объекта
Одним из широко используемых токсикогентических методов является анализ хромосомных аберраций в меристематической ткани проростков корешков Allium cepa [4]. Allium cepa хорошо изучен генетически, имеет 16 (2n = 16) удобных для анализа хромосом. Материал – луковицы можно получать в достаточно большом количестве в любое время года. Метод является хорошо разработанным и информативным. На одном и том же материале можно характеризовать изучаемый фактор среды по трем параметрам:
–мутагенному действию;
–митотоксическому действию;
–судить о причинах нарушения митотического цикла.
5.3.1.Клеточный (митотический) цикл
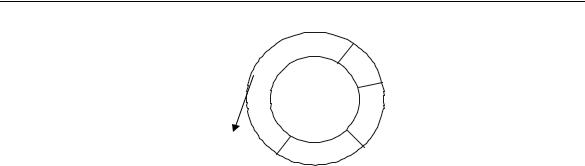
Клеточный (митотический) цикл – это совокупность взаимосвязанных и хронологически детерминированных событий, происходящих в клетке в период подготовки к делению, а также на протяжении самого митоза (рис. 5.3.1).
71
Генетическая токсикология
G1
G0
2 n 2
c
|
М |
|
и |
|
т |
|
о |
|
з |
|
2 |
|
n |
|
4 |
|
G |
2 |
c |
2 |
|
c4n |
|
S
Рис. 5.3.1 Клеточный (митотический цикл)
Таким образом, митотический цикл совпадает со всем периодом существования клетки, то есть с её жизненным циклом. В митотическом цикле выделяют четыре периода: собственно митоз, пресинтетический период, синтетический и постсинтетический. Три последних периода приходятся на интерфазу. Митотической цикл может быть изображен в виде круга, на котором, пропорционально продолжительности, отмечены его периоды от одного митоза до другого (рис. 5.3.2).
Интерфазой называют стадию покоящегося ядра, однако это наиболее активная фаза с точки зрения биохимических процессов. В интерфазной клетке хорошо видно ядро. Оно имеет гомогенную мелкозернистую структуру с нанесенным рисунком хроматина, хорошо видны ядрышки (рис. 5.3.2). В пресинтетический период (G1) в клетке происходит активный синтез белков, обеспечивающих рост клетки, накопление продуктов, необходимых для редупликации ДНК. Этот период наиболее длительный (от 10 часов до нескольких суток). Генетическая характеристика клеток в этот период 2n2c.
В синтетический период (S) проходит редупликация ДНК, наработка гистонов. Продолжительность его 6 – 10 часов. Генетическая характеристика 2n4c. Хромосомы удваиваются, то есть в каждой хромосоме теперь две молекулы ДНК, упакованные в две хроматиды (рис. 5.3.3).
В постсинтетический период (G2) заканчивается подготовка клетки к делению. Идут подготовительные процессы для формирования ахроматинового веретена, идет накопление энергии, поскольку митоз требует значительных затрат энергии. Генетическая характеристика клеток в этот период 2n4c.
72
Глава 5. Методы выявления и оценки генотоксикантов
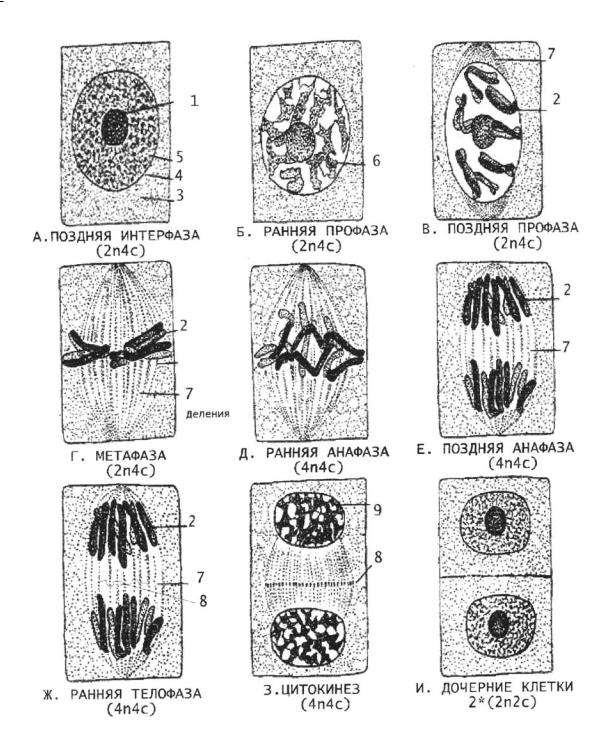
Рис. 5.3.2. Митотическое деление растительной клетки: 1 – ядро, 2 – хромосомы, 3 – цитоплазма, 4 – кариолемма, 5 – кариоплазма, 6 – глыбки хромтина, 7 – веретено деления, 8 – фрагмопласт, 9 – ядра дочерних клеток
73

Генетическая токсикология
G1 S G2
Дочерние
клетки
Рис. 5.3.3. Морфология хромосом в течение клеточного цикла
Интерфаза от G1 и до конца S периода называется ранней интерфазой. Её генетическая характеристика 2n2c. Хромосома в этот период содержит одну молекулу ДНК. От конца S периода и до митоза – поздняя интерфаза. Каждая хромосома содержит по две молекулы ДНК, упакованные в две хроматиды. Генетическая характеристика этого периода клеточного цикла 2n4c.
Далее клетка вступает в митоз. Количество митозов, которое может пройти клетка, генетически детерминировано. Использовав определенное количество митозов, клетка может перейти в особую фазу митотического цикла – G0 или дифференцированное состояние. В клетке могут идти активные метаболические процессы, но в митоз она не вступает. Разные клетки в разное время вступают в G0. Например, нервные клетки рано перестают делиться, клетки кожного эпителия длительное время сохраняют способность к делению.
Митоз. Картины различных фаз митоза в настоящее время хорошо изучены.
Профаза. Начало митоза связано с заметным увеличением размеров ядра и появлением в нем хромосомных нитей. Это обусловлено спирализацией и дегидратацией хромосом. За счет многопорядковой спирализации хромосомы и становятся видимыми в световой микроскоп (рис. 5.3.2). Для профазы характерно специфическое поведение клеточного центра. Центриоли удваиваются еще в интерфазу. В профазу удвоенные центриоли начинают отходить друг от друга к противоположным полюсам клетки. Между ними натягиваются полимеризованные белковые нити – образуется ахроматиновое веретено деления (рис. 5.3.2). В конце
74