

7.) Особенности fish метода.
Флуоресцентная гибридизация на препарате in siti FISH - Fluorescent In Situ Hybrization включает применение специально подготовленных (флюоресцирующих) ДНК проб.
ДНК – зонды представляют собой клонированные фрагменты генома человека для выявления генетических дефектов на хромосомном уровне.
При этом используется многоцветовая флуоресцентная in siti гибридизация ДНК на препарате метафазных хромосом или интерфазных клеток для маркировки хромосом или их участков. Данная диагностика, охватывает практически весь спектр хромосомных аномалий, что в первую очередь позволяет выделять редкие хромосомные синдромы. Метод FISH может применяться и для диагностики анеуплоидий в интерфазных ядрах.
8.) Мутации, выявляемые цитогенетическим методом (геномные и хромосомные), причины и механизмы возникновения.
Причиной структурных изменений генома может быть нарушение тех процессов, которые в норме обеспечивают его устойчивость, в первую очередь процессов, протекающих в мейозе.
Так, нарушение кроссинговера, приводящее к обмену неравноценными участками ДНК между хроматидами, может привести к утрате или удвоению определенной нуклеотидной последовательности в них. Если это затрагивает структуру отдельного гена, то возможно возникновение генной мутации с изменением количества нуклеотидов в нем (см. разд. 3.4.2.3). Если при неравноценном обмене затронут участок хроматиды, содержащий несколько генов, изменяется доза этих генов в геноме. Он либо лишается каких-то генов (деления), либо эти гены оказываются в геноме в двойном количестве (дупликапия). Изменение дозового соотношения отдельных генов наблюдается также при разных видах хромосомных перестроек, не обязательно связанных с неравноценным кроссинговером (см. разд. 3.5.3.3).
Нарушение расхождения бивалентов в анафазе I мейоза является причиной изменения количества хромосом в гаплоидном наборе гамет. Нерасхождение отдельного бивалента приводит к появлению одной гаметы, лишенной данной хромосомы, и другой, имеющей эту группу сцепления в двойном количестве (рис. 3.76). Оплодотворение таких гамет нормальными половыми клетками приводит к появлению особей, в кариотипе которых изменено общее число хромосом за счет уменьшения (моносомия) или увеличения (трисомия) числа отдельных хромосом. Нарушения структуры генома, заключающиеся в изменении количества отдельных хромосом, называют анэуплоидией.
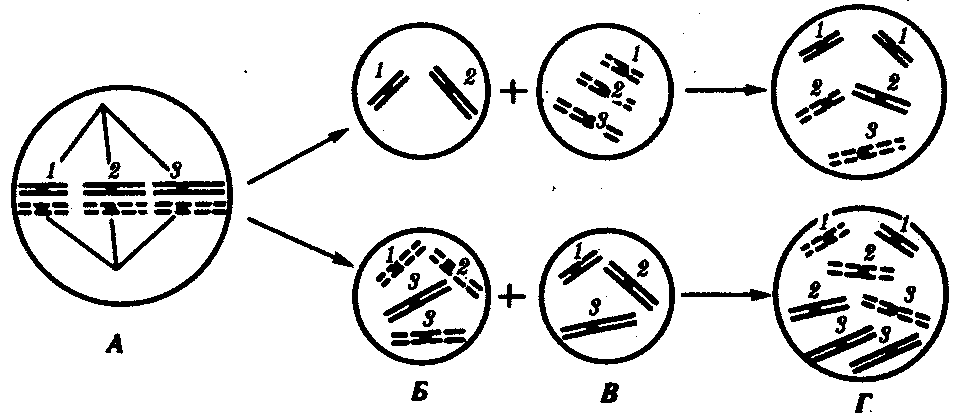
Рис. 3.76. Нарушение расхождения отдельных бивалентов (1, 2, 3) в мейозе
как причина возникновения анэуплоидий:
А — метафаза 1 мейоза; Б — образование аномальных гамет в результате нарушения расхождения 3-го бивалента в анафазе I мейоза; В — оплодотворение аномальных гамет нормальными гаметами другого пола; Г — образование зигот с анэуплоидным кариотипом (моносомия или трисомия по 3-й хромосоме, соответственно сверху и снизу)
В том случае, если в целом повреждается механизм распределения гомологичных хромосом между полюсами веретена (что наблюдается при его разрушении), клетка остается неразделившейся. Во второе деление мейоза она вступает не гаплоидной, а диплоидной. Из нее образуются диплоидные гаметы. Оплодотворение таких гамет приводит к образованию триплоидных организмов. Увеличение в кариотипе зиготы числа наборов хромосом называют полиплоидией
В основе изменения структуры хромосомы, как правило, лежит первоначальное нарушение ее целостности — разрывы, которые сопровождаются различными перестройками, называемыми хромосомными мутациями или аберрациями.
Разрывы хромосом происходят закономерно в ходе кроссинговера, когда они сопровождаются обменом соответствующими участками между гомологами (см. разд. 3.6.2.3). Нарушение кроссинговера, при котором хромосомы обмениваются неравноценным генетическим материалом, приводит к появлению новых групп сцепления, где отдельные участки выпадают — делении — или удваиваются — дупликации (рис. 3.57). При таких перестройках изменяется число генов в группе сцепления.
Разрывы хромосом могут возникать также под влиянием различных мутагенных факторов, главным образом физических (ионизирующего и других видов излучения), некоторых химических соединений, вирусов.
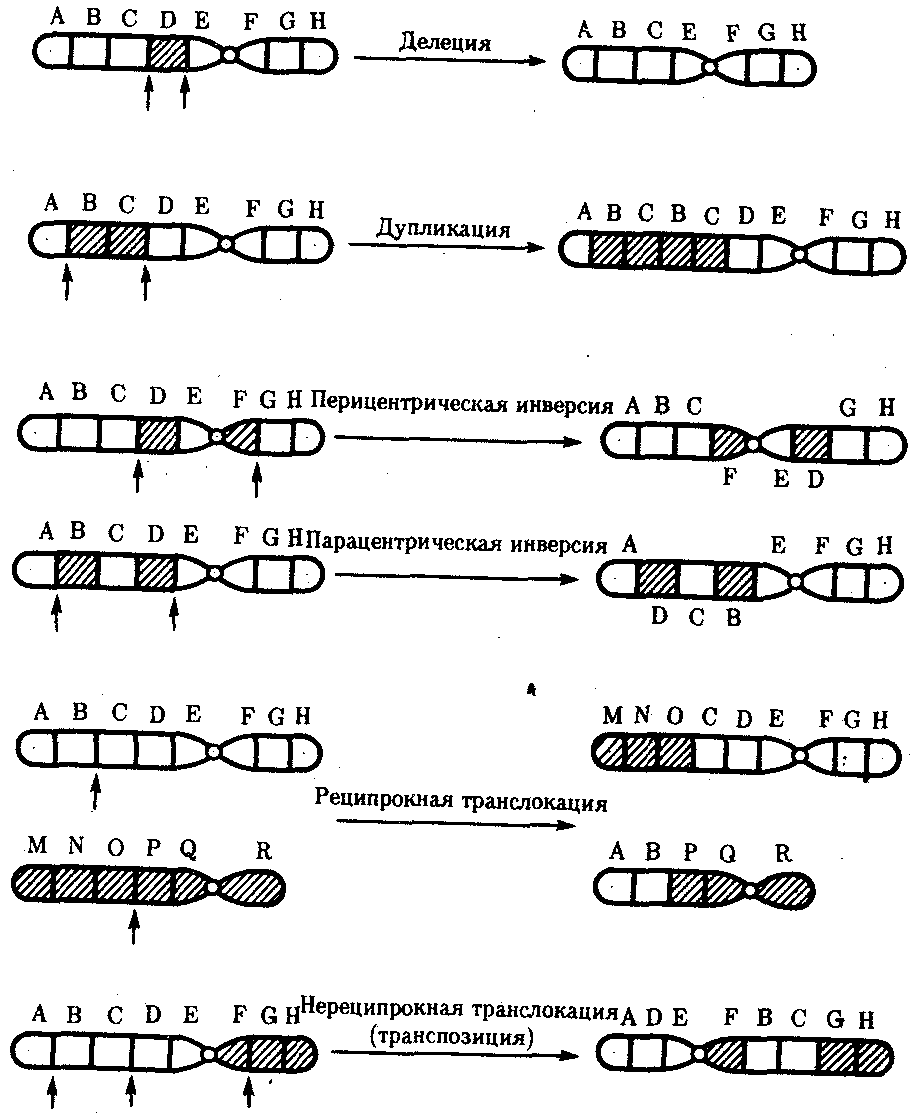
Рис. 3.57. Виды хромосомных перестроек
Нарушение целостности хромосомы может сопровождаться поворотом ее участка, находящегося между двумя разрывами, на 180° — инверсия. В зависимости от того, включает ли данный участок область центромеры или нет, различают перицентрические и парацентрические инверсии (рис. 3.57).
Фрагмент хромосомы, отделившийся от нее при разрыве, может быть утрачен клеткой при очередном митозе, если он не имеет центромеры. Чаще такой фрагмент прикрепляется к одной из хромосом — транслокация. Нередко две поврежденные негомологичные хромосомы взаимно обмениваются оторвавшимися участками —ре-ципрокная транслокация (рис. 3.57). Возможно присоединение фрагмента к своей же хромосоме, но в новом месте — транспозиция (рис. 3.57). Таким образом, различные виды инверсий и транслокаций характеризуются изменением локализации генов.
Хромосомные перестройки, как правило, проявляются в изменении морфологии хромосом, что можно наблюдать в световой микроскоп. Метацентрические хромосомы превращаются в субметацентрические и акроцентрические и наоборот (рис. 3.58), появляются кольцевые и полицентрические хромосомы (рис. 3.59). Особую категорию хромосомных мутаций представляют аберрации, связанные с центрическим слиянием или разделением хромосом, когда две негомологичные структуры объединяются в одну — робертсоновская транслокация, или одна хромосома образует две самостоятельные хромосомы (рис. 3.60). При таких мутациях не только появляются хромосомы с новой морфологией, но и изменяется их количество в кариотипе.
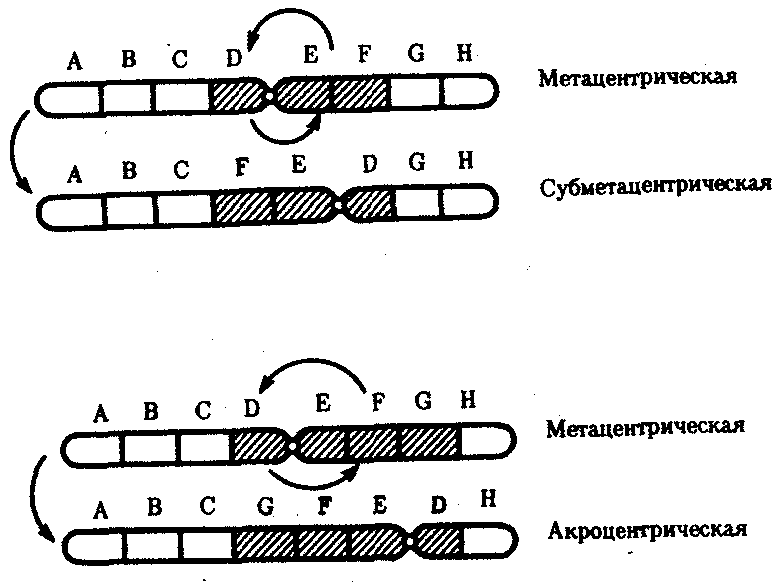
Рис. 3.58. Изменение формы хромосом
в результате перицентрических инверсий
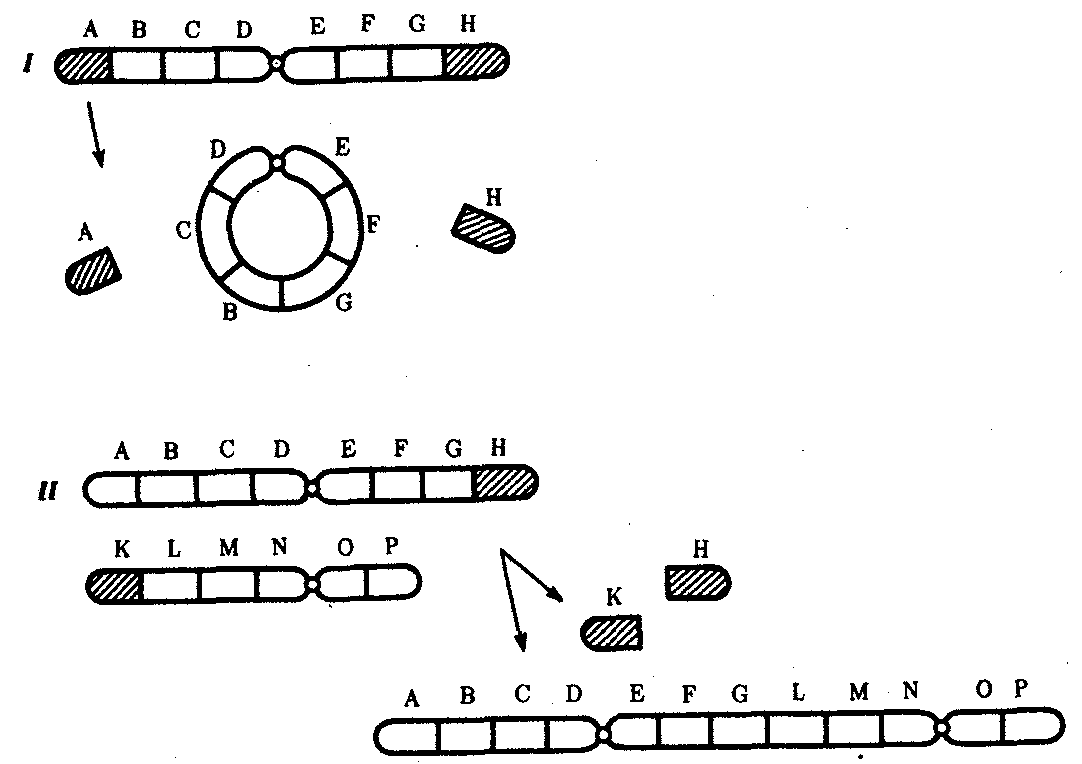
Рис. 3.59. Образование кольцевых (I) и полицентрических (II) хромосом
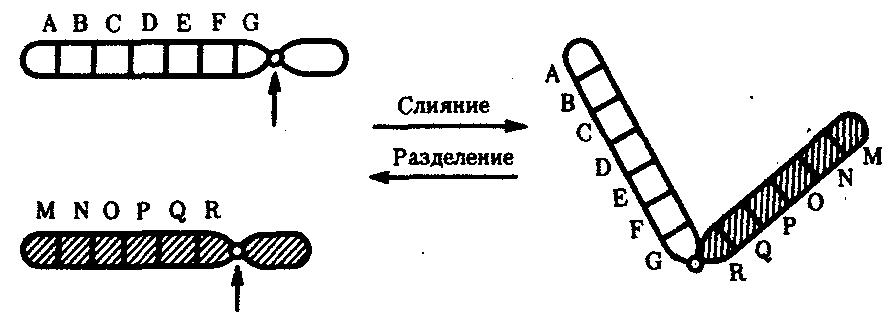
Рис. 3.60. Хромосомные перестройки, связанные с центрическим слиянием
или разделением хромосом являются причиной изменения числа хромосом
в кариотипе