

Гидролиз эфиров холестерина
Эфиры холестерина (ЭХС) расщепляются на свободный холестерин и высшие жирные кислоты (ВЖК) под действием фермента – холестеролэстеразы панкреатического и кишечного соков:
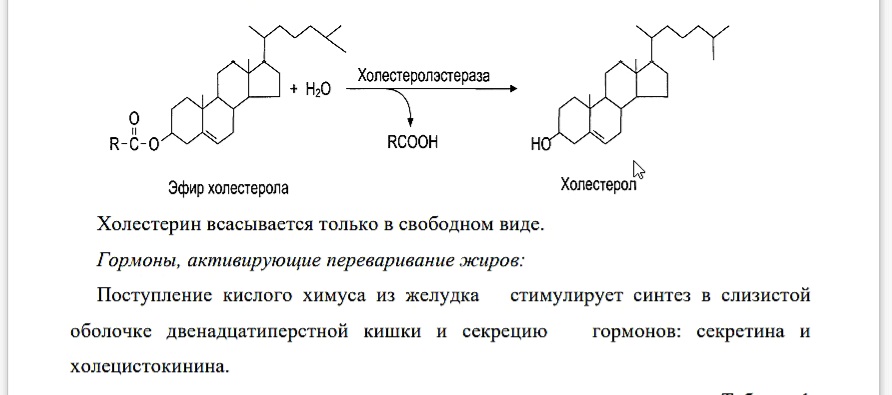
Холестерин всасывается только в свободном виде.
Образование смешанной мицеллы и всасывание продуктов переваривания липидов Плохо растворимые в водной среде, продукты гидролиза липидов: высшие жирные кислоты, 2-МАГ, холестерол, а также поступившие с пищей жирорастворимые витамины А, Д, Е, К включаются в мицеллы желчи, образуя смешанные мицеллы (рис. 1-5).
Мицелла – это сферический комплекс, в центре которого находятся транспортируемые гидрофобные продукты переваривания, окруженные желчными кислотами. В состав гидрофобного ядра мицеллы входят: ацильные остатки жирных кислот, 2-моноацилглицеролов, неполярные группы холестерола и желчных кислот. Большую роль в формировании мицелл играют молекулы лизофосфатидной кислоты (продукт гидролиза фосфолипидов) и желчные кислоты, которые с помощью своих отрицательно заряженных групп образуют гидрофильную оболочку. Желчные кислоты и их соли, стабилизируют смешанные мицеллы, способствуя их всасыванию в клетки кишечника.
Смешанные мицеллы:
• растворимы в водной фазе содержимого тонкой кишки
• имеют малые размеры: в 100 раз меньше самых мелких эмульгированных жировых капель.
Образование смешанной мицеллы Мицеллы перемещаются к всасывающей поверхности кишечного эпителия, а затем, внутри клеток распадаются на составные компоненты.
Проникновение мицелл в энтероцит регулируется гликокаликсом, покрывающим апикальную мембрану энтероцитов. Гликокаликс работает избирательно: отбрасывает назад в полость крупные гидрофобные молекулы и пропускает мелкие гидрофильные. Продукты ферментативного гидролиза липидов, которые хорошо растворимы в воде, такие как: глицерин, аминоспирты, фосфорная кислота, жирные кислоты с короткой углеродной цепью (менее 10 атомов углерода) всасываются в кишечнике, попадая в энтероцит, а откуда поступают в кровь воротной вены и доставляются в печень. Всосавшиеся продукты гидролиза в энтероцитах используются для синтеза липидов, близких по составу к липидам организма.
Желчные кислоты, оставшиеся в просвете кишечника и не вошедшие в состав мицелл, выводятся с калом (около 5%). Потери желчных кислот в кишечнике (1-1,5 г) восполняются за счет их синтеза в печени из холестерола. Основная масса желчных кислот (90-95%), поступивших из мицелл в энтероциты, всасывается и с кровью по воротной вене возвращаются обратно в печень. Откуда, через желчные протоки, они вновь поступают в желчный пузырь и затем, в составе простой мицеллы желчи, изливаются в кишечник. В этом случае имеет место постоянная циркуляция желчных кислот между печенью и кишечником. Данный процесс получил название энтерогепатической циркуляции желчных кислот (рис.1-6). Рис.1-6. Энтерогепатическая циркуляция Установлено, что у человека общий пул желчных кислот составляет примерно 2,8–3,5 г, при этом они совершают 6–8 оборотов в сутки, (в зависимости от количества жира в пище).
Биологическое значение энтерогепатической циркуляции: для эмульгирования липидов требуется меньшее количество желчных кислот. I-й ресинтез липидов в энтероцитах и его биологическое значение.
Процесс синтеза жира в энтероцитах из продуктов гидролиза липидов, называется I-м ресинтезом жира.
Ресинтез ТАГ в энтероцитах протекает в гладком эндоплазматическом ретикулуме из 2-моноацилглицеролов и ВЖК. В ресинтезе жиров участвуют не только жирные кислоты, всосавшиеся из кишечника, но и жирные кислоты, синтезированные в организме, поэтому по составу ресинтезированные жиры отличаются от жиров, полученных с пищей.
Первая стадия ресинтеза жиров - реакция активации жирных кислот. Жирные кислоты активируются при участии ацил КоА-синтетаз, специфичных к длине углеводородного радикала:

Вторая стадия ресинтеза жиров - ацилирование 2-МАГ до ТАГ активированными жирными кислотами при участии ферментов — трансацилаз (или ацилтрансфераз): Ресинтез фосфолипидов в энтероцитах Большая часть фосфолипидов содержимого тонкой кишки приходится на фосфатидилхолин (лецитин), поэтому в энтероцитах преимущественно этерифицируется именно лизофосфатидилхолин: Ресинтез эфиров холестерина В микросомах клеток слизистой оболочки тонкой кишки, всосавшиеся молекулы ХС превращаются в эфиры, путём взаимодействия с ацил-KоA при участии фермента ацилхолестеролацилтрансферазы (АХАТ). От активности данного фермента зависит скорость поступления экзогенного ХС в организм
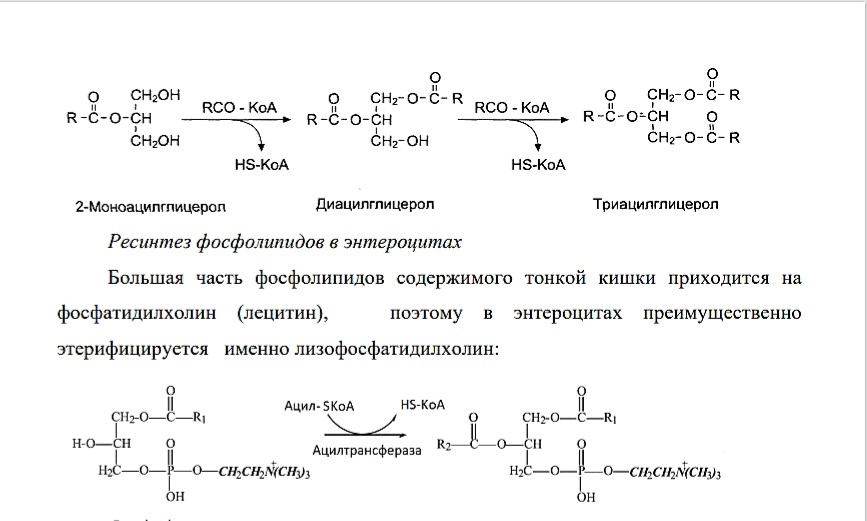
Биологическое значение ресинтеза липидов заключается в том, что синтезируются липиды, специфичные для организма человека. Однако возможности «адаптировать» в процессе ресинтеза состав пищевых жиров к составу жиров организма человека ограниченны.