

Роль фосфорилазы при мобилизации гликогена
2. α(1,4)-α(1,4)-Глюкантрансфераза – фермент, переносящий фрагмент из трех остатков глюкозы на другую цепь с образованием новой α1,4-гликозидной связи. При этом на прежнем месте остается один остаток глюкозы и "открытая" доступная α1,6-гликозидная связь.
3. Амило-α1,6-глюкозидаза, ("деветвящий" фермент) – гидролизует α1,6-гликозидную связь с высвобождением свободной (нефосфорилированной) глюкозы. В результате образуется цепь без ветвлений, вновь служащая субстратом для фосфорилазы.
Гликолиз: общая характеристика, стадии, реакции процесса, регулируемые ферменты, энергетический эффект. Судьба про- дуктов гликолиза в аэробных условиях: схема процесса, связь с синтезом АТФ.
Путь, по которому глюкоза окисляется до пировиноградной кислоты для получе-
ния энергии, называется гликолизом. В зависимости от дальнейшей судьбы пирувата различают аэробный и анаэробный гликолиз.
В аэробном процессе пировиноградная кислота превращается в ацетил-S-КоА и
далее сгорает в реакциях тканевого дыхания до СОB2B и НB2BО.
В анаэробном процессе пировиноградная кислота восстанавливается до молоч-ной кислоты (лактата), поэтому в микробиологии анаэробный гликолиз называют
молочнокислым брожением. Лактат является метаболическим тупиком и далее ни во что не превращается, единственная возможность утилизовать лактат – это окислить его
обратно в пируват. Практически все клетки организма способ-ны к анаэробному гликолизу. Для эритроцитов
он является единственным источником энергии. Клетки скелетной мускулатуры за счет бескислородного расщепления глюкозы способны выполнять мощную, быструю, интенсивную работу, как, например, бег на короткие дистанции, напряжение в силовых видах спор-
та.
А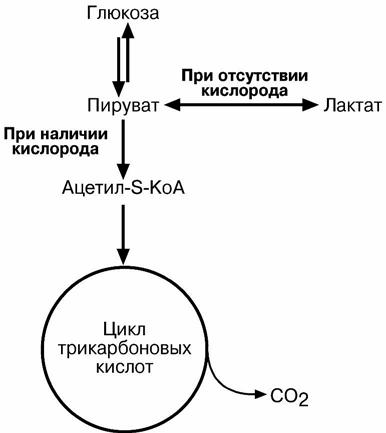 наэробный
гликолиз локализуется в цито-золе и
включает 2 этапа из 11 ферментатив-ных
реакций
наэробный
гликолиз локализуется в цито-золе и
включает 2 этапа из 11 ферментатив-ных
реакций
Первый этап – подготовительный, здесь происходит
з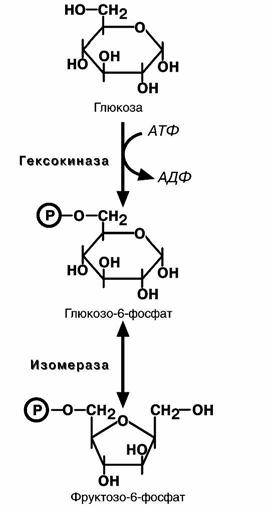 атрата
энергии АТФ, активация глюкозы и
образование
атрата
энергии АТФ, активация глюкозы и
образование
из нее триозофосфатов.
Первая реакция гликолиза, строго говоря, к гликоли-
зу не относится. Это гексокиназная реакция о которой
ранее уже говорилось ("Метаболизм гликогена"). Ее роль сводится к превращению глюкозы в реакционно способное соединение за счет фосфорилирования 6-го, невключенного в кольцо, атома углерода.
Для печени характерен изофермент гексокиназы –
глюкокиназа. Низкое сродство этого фермента к глюкозе обеспечивает ее захват печенью только после приема пищи, когда создается высокая концентрация глюкозы в
крови.
При обычных концентрациях глюкозы в крови печень ее не потребляет и та достается другим тканям.
Вторая реакция – реакция изомеризации – необхо-
дима для выведения еще одного атома углерода из
кольца для его последующего фосфорилирования. В ней образуется фруктозо-6-фосфат.
Т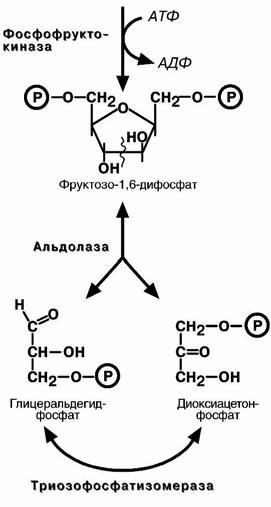 ретья
реакция –
фосфорилирование
фруктозо-6-
ретья
реакция –
фосфорилирование
фруктозо-6-
фосфата с образованием почти симметричной молекулы фруктозо-1,6-дифосфата.
В четвертой реакции фруктозо1,6-дифосфат разрез
ается пополам с образование двух фосфорилирован-ных триоз-изомеров, альдозы глицеральдегида (ГАФ) и
кетозы диоксиацетона (ДАФ).
Пятая реакция подготовительного этапа – переход глицеральдегидфосфата и диоксиацетонфосфата друг в друга. Равновесие реакции сдвинуто в пользу диоксиацетонфосфата, его доля составляет 97%, доля глицеральдегидфосфата – 3%. Эта реакция, при всей ее простоте, является вершителем судьбы глюкозы:
при нехватке энергии в клетке и активации окисления глюкозы диоксиацетонфосфат превращается в глицеральдегидфосфат, который далее окисляется на втором этапе гликолиза;
при достаточном количестве АТФ, наоборот, глицеральдегидфосфат изомеризуется в диоксиацетонфосфат, и последний отправляется на синтез жиров (см "Синтез триацилглицеридов").
Второй этап гликолиза – это освобождение
э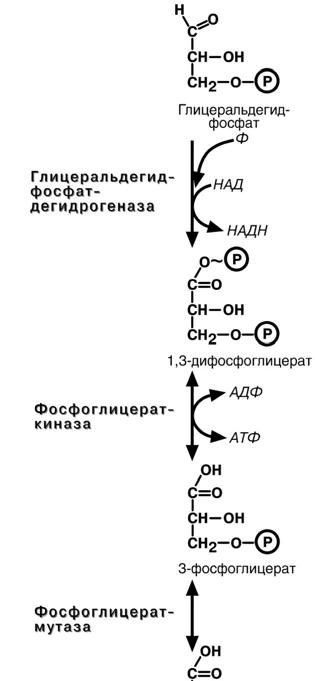 нергии,
содержащейся в глицеральдегидфосфате,
нергии,
содержащейся в глицеральдегидфосфате,
и запасание ее в форме АТФ.
Шестая реакция гликолиза – окисление глице-
ральдегидфосфата и присоединение к нему фос-
форной кислоты приводит к образованию макроэр-гического соединения 1.3-дифосфоглицериновой
кислоты.
В седьмой реакции энергия фосфоэфирной связи, заключенная в 1,3-дифосфоглицерате тра-
тится на образование АТФ. Реакция получила до-
полнительное название –реакция субстратного фосфорилирования, что уточняет источник энергии
для получения макроэргической связи в АТФ (суб-
страт) в отличие от окислительного фосфорилиро-вания (электрохимический градиент ионов водоро-
да на мембране митохондрий).
Подобных реакций в клетке всего три – 1) ука-занная реакция, 2) пируваткиназная реакция, деся-тая реакция гликолиза (см ниже), 3) тиокиназная реакция цикла трикарбоновых кислот.
Восьмая реакция – синтезированный в преды-
дущей реакции 3-фосфоглицерат изомеризуется в
2-фосфоглицерат.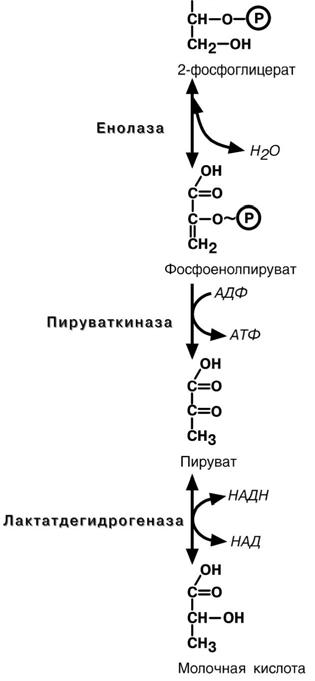
Девятая реакция – отрыв молекулы воды от
2-фосфоглицериновой кислоты приводит к образо-
ванию еще одной макроэргической фосфоэфирной связи.
Еще одна реакция субстратного фосфорили-рования – десятая реакция гликолиза – заключается в переносе макроэргического фосфата с фосфоенолпирувата на АДФ.
Последняя реакция , одиннадцатая, образова-ние молочной кислоты из пирувата под действием
лактатдегидрогеназы. Важно то, что эта реакция
осуществляется только в анаэробных условиях. Эта реакция необходима клетке, так как НАДН, об-
разующийся в 6-й реакции, в отсутствие кислорода не может поступать и окисляться в митохондриях.
При наличии кислорода пировиноградная кислота превращается в ацетил-S-КоА.
При аэробном гликолизе из 1 молекулы глюкозы образуется 36 или 38 АТФ, выделяется 6 СО2 и 6 Н2О.
Анаэробный распад глюкозы (анаэробный гликолиз). Судьба продуктов гликолиза в анаэробных условиях. Биологическое значение анаэробного распада глюкозы.
Анаэробный гликолиз, или фосфотриозный путь, или шунт Эмбдена-Мейерхофа включает 10 реакций. Ступенчатое окисление глюкозы создает возможность не только преодоления при обычной температуре порога энергии активации отдельных реакций, но и рационального использования энергии, освобождающейся здесь не в форме взрыва, а шаг, за шагом, отдельными порциями. На основе гликолиза возникло кислородное дыхание. Гликолиз – это древнее усилие природы использовать энергию – студенты пишут отдельно реакции формулами, а пояснение к реакциям в тексте лекции (как ЦТК).
1 реакция - глюкоза фосфорилируется под действием фермента глюкокиназы (в печени) или гексокиназы (в других тканях)
2 реакция – глюкозо-6-фосфат изомеризуется во фруктозо-6-фосфат под действием фосфогексоизомеразы; этот фермент действует на молекулу с открытой линейной конфигурацией
3 реакция - фруктозо-6-фосфат необратимо фосфорилируется во фруктозо-1,6-дифосфат под действием фосфофруктокиназы, которая также действует на молекулу с открытой конфигурацией
4 реакция – фруктозо-1,6-дифосфат под действием альдолазы расщепляется на 2 фосфотриозы – ФГА и ДОАФ, реакция обратимая
ДОАФ может участвовать в синтезе ТАГ и ФЛ, восстанавливаясь до глицерофосфата, также участвует в глицерофосфатном челночном механизме, но основная его масса переходит в ФГА
5 реакция – ФГА окисляется с участием НАД и фосфорилируется. При этом энергия окисления трансформируется в макроэргическую связь 1,3 дифосфоглицерата.
6 реакция – 1,3-дифосфоглицерат реагирует с АДФ, отдает ей остаток фосфорной кислоты и выделяется АТФ. Так происходит субстратное фосфорилирование и образуется 3-фосфоглицерат, реакцию ускоряет фосфоглицераткиназа
7 реакция – под влиянием фосфоглицеромутазы остаток фосфорной кислоты переносится с С3 на С2 и образуется 2-фосфоглицерат
8 реакция – 2-фосфоглицерат дегидратируется енолазой. При этом за счет внутримолекулярной ОВР энергия аккумулируется в виде макроэргической связи в фосфоенолпирувате. Енолаза ингибируется ионами фторида; этим пользуются в тех случаях, когда необходимо остановить гликолиз, например, перед определением содержания глюкозы в крови. Енолаза нуждается в ионах магния и марганца:
9 реакция – ФЕП передает остаток фосфорной кислоты на АДФ, при этом образуется енолпируват и выделяется АТФ, вновь происходит субстратное фосфорилирование. Реакция ускоряется пируваткиназой. Енолпируват спонтанно превращается в ПВК.
10 реакция - ПВК в анаэробных условиях восстанавливается в молочную кислоту (лактат)
Энергетический баланс анаэробного гликолитического окисления глюкозы
Если процесс гликолиза начинается с глюкозы, то на образование фруктозо-6-фосфата и фруктозо-1,6-дифосфата затрачивается 2 молекулы АТФ. Т.к., в результате гликолиза образуется 4 АТФ, следовательно, в чистом виде запасается 2 АТФ. Если процесс гликолиза начинается с глюкозо-6-фосфата, образованного при распаде гликогена, затрачивается 1 АТФ для образования фруктозо-1,6-дифосфата, тогда выделяется 3 АТФ.
При анаэробном гликолизе из 1 молекулы глюкозы образуется 2 молекулы лактата и 2 АТФ.
Пентозофосфатный цикл осуществляется в анаэробных условиях, в результате этого процесса выделяется 6СО2 и 12НАДФН2. НАДФН2 необходимы для восстановительного синтеза.
Биосинтез глюкозы (глюконеогенез). Субстраты, энергетиче- ские затраты, регулируемые ферменты. Цикл Кори.
Глюконеогенез – это синтез глюкозы из неуглеводных компонентов: лактата, пирувата. глицерола, метаболитов цикла Кребса, аминокислот. Все аминокислоты,
кроме кетогенных лейцина и лизина, способны участвовать в синтезе глюкозы. Некоторые из них – глюкогенные – полностью включаются в молекулу глюкозы, некоторые – смешанные – частично.
организме всегда существует потребность в глюкозе:
для эритроцитов глюкоза является единственным источником энергии,
нервная ткань потребляет 120 г глюкозы в сутки, притом эта величина не за-висит от интенсивности ее работы. Только в экстремальных ситуациях (дли-тельное голодание) она способна получать энергию из неуглеводных источников,
глюкоза играет весомую роль для поддержания необходимых концентраций метаболитов цикла трикарбоновых кислот (в первую очередь оксалоацетата).
Таким образом, при определенных ситуациях – при низком содержании углеводов в пище, голодании, длительной физической работе – организм должен иметь возможность получить глюкозу. Это достигается процессом глюконеогенеза. Кроме получения глюкозы, глюконеогенез обеспечивает и уборку "шлаков" – лактата, образованного при мышечной работе и в эритроцитах, и глицерола, являющегося продуктом липолиза в жировой ткани.
Глюконеогенез лишь отчасти повторяет реакции окисления глюкозы. Как указывалось ранее, в гликолизе существуют три необратимые стадии: пируваткиназная(десятая), фосфофруктокиназная (третья) и гексокиназная (первая). На этих стадиях существуют энергетические барьеры, которые обходятся с помощью специальных реакций.
Обход десятой реакции гликолиза
На этом этапе глюконеогенеза работают два основных фермента – в митохондриях пируваткарбоксилаза и в цитозоле фосфоенолпируват-карбоксикиназа.
Пируваткарбоксилаза превращает пировиноградную кислоту в оксалоацетат. Необ-ходимо отметить, что эта реакция идет в клетке постоянно, являясь анаплеротиче-
ской
(пополняюшей) реакцией ЦТК. Далее
оксалоацетат должен попасть в цитозоль
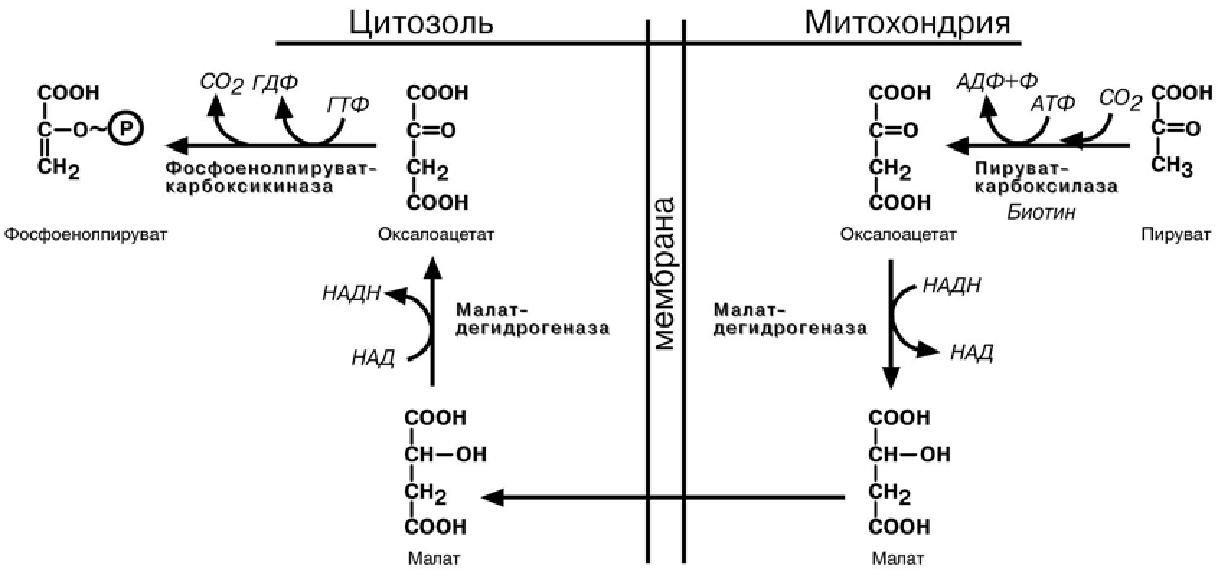 и
превратиться в фосфоенолпируват. Однако
дело осложняется непроницаемостью
мембраны для оксалоацетата. Зато через
мембрану может пройти малат, предшест-венник
оксалоацетата по ЦТК. Так как в условиях
недостаточности глюкозы в клетке
активируется липолиз и окисление жирных
кислот, то повышается количество НАДН
в митохондриях. Этот избыток позволяет
повернуть малатдегидрогеназную реакцию
ЦТК вспять. Малат накапливается, выходит
в цитозоль и здесь превращается в
оксалоацетат.
и
превратиться в фосфоенолпируват. Однако
дело осложняется непроницаемостью
мембраны для оксалоацетата. Зато через
мембрану может пройти малат, предшест-венник
оксалоацетата по ЦТК. Так как в условиях
недостаточности глюкозы в клетке
активируется липолиз и окисление жирных
кислот, то повышается количество НАДН
в митохондриях. Этот избыток позволяет
повернуть малатдегидрогеназную реакцию
ЦТК вспять. Малат накапливается, выходит
в цитозоль и здесь превращается в
оксалоацетат.
В цитоплазме фосфоенолпируват-карбоксикиназа
осуществляет превращение оксалоацетата в фосфоенолпируват, для реакции требуется энергия ГТФ. От молекулы отщепляется тот же углерод, что и присоединяется.
Обход третьей реакции гликолиза
В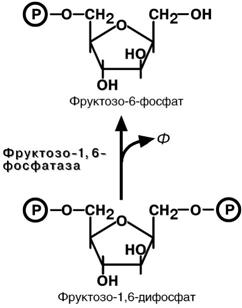 торое
препятствие на пути синтеза глюкозы –
фос-
торое
препятствие на пути синтеза глюкозы –
фос-
фофруктокиназная реакция – преодолевается с помощью
фермента
фруктозо-1,6-дифосфатазы. Этот фермент
есть в почках, печени, поперечно-полосатых
мышцах. Таким образом, эти ткани способны
синтезировать фруктозо-6-фосфат и
глюкозо-6-фосфат.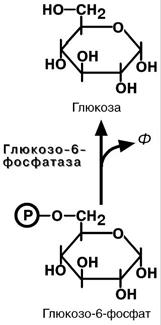
Обход первой реакции гликолиза
Последняя реакция катализируется глюкозо-6-
фосфатазой . Она имеется только в печени и почках, следовательно, только эти ткани могут продуцировать свободную глюкозу.
ГЛЮКОЗО- ЛАКТАТНЫЙ И ГЛЮКОЗО- АЛАНИНОВЫЙ ЦИКЛЫ( ЦИКЛ КОРИ)
Глюкозо-лактатный цикл (цикл Кори) – это связь глюконеогенеза в печени и образования лактата эритроцитах или мыш-цах из глюкозы. В эритроцитах молочная
кислота образуется
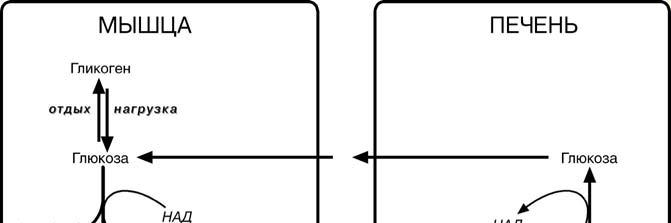
непрерывно, так как для них анаэробный
г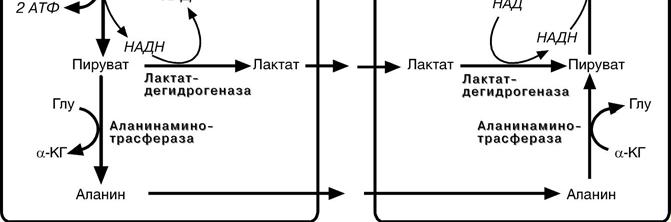 ликолиз является
ликолиз является
единственным спосо-бом образования энер-
гии. В скелетных мыш-цах накопление лакта-та является следстви-ем гликолиза при очень интенсивной, максимальной мощно-
сти, работе, и чем более такая работа интенсивна, тем менее продолжительна По-
сле нагрузки (во время восстановления) лактат удаляется из мышцы довольно бы-
стро – всего за 0,5-1,5 часа.
Дополнение
Следует
отметить, что если продолжи-
тельность нагрузки мала (до 10 секунд),
то количество АТФ пополняется пре-имущественно в
креатинфосфоки-назной
реакции. В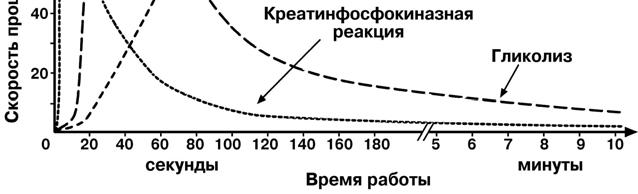
таком режиме к при-меру работают мышцы у ттанги-
стов, прыгунов как в длину, так и в высо-
ту, метателей мо-лота, копья и т.п..
Если нагрузка не бо-лее 90 секунд – АТФ синтезируется в основном в реакциях анаэробного глико-
лиза. В спорте это бегуны-спринтеры на 100-500 м, спортсмены силовых ви-дов (борцы, тяжелоатлеты, бодибилдеры). Если напряжение мышцы длится
более двух минут – развивается аэробное окисление глюкозы в реакциях ЦТК
и дыхательной цепи.
Но, хотя мы и говорим об аэробном окислении глюкозы, необходимо знать и помнить, что лактат образуется в мышце всегда: и при анаэробной, и при аэробной работе, однако в разных количествах.
Образовавшийся лактат может утилизоваться только одним способом – превратиться в пировиноградную кислоту. Но, как уже указывалось, пируват токсичен для клеток и должен быть как можно быстрее утилизован. Сама мышца ни при работе, ни во время отдыха не занимается превращением лактата в пируват из-за наличия специфического изофермента ЛДГ-5.
Если молочная кислота поступила в миокардиоциты, она быстро превраща-ется в пируват, далее в ацетил-S-КоА и вовлекается в полное окисление до
СОB2B и НB2BО.
Большая часть лактата захватывается гепатоцитами, окисляется в пировино-
градную кислоту и вступает на путь глюконеогенеза.
Целью глюкозо-аланинового цикла также является уборка пирувата, но, кроме этого решается еще одна немаловажная задача – уборка лишнего азота из мышцы.
При мышечной работе и в покое в миоците распадаются белки и образуемые амино-кислоты рансаминируются с α-кетоглутаратом. Полученный глутамат взаимодействует с пируватом. Образующийся аланин является транспортной формой и пирувата и азота из мышцы в печень. В гепатоците идет обратная реакция трансаминирова-ния, аминогруппа передается на синтез мочевины, пируват используется для синте-за глюкозы
Глюкоза, образованная в печени из лактата или аланина, возвращается обратно в мышцы, восстанавливая во время отдыха запасы гликогена.
Кроме мышечной работы, глюкозо-аланиновый цикл активируется во время голодания, когда мышечные белки распадаются и многие аминокислоты используются
качестве источника энергии, а их азот необходимо доставить в печень.
7. Пентозо-фосфатный путь (ПФП) окисления глюкозы. Биоло- гическое значение.
Пентозофосфатный путь окисления глюкозы не связан с образованием энергии.
Значение ПФП:
Образование НАДФН
для синтеза жирных кислот,
холестерина и других стероидов,
для синтеза глутаминовой кислоты из α-кетоглутаровой кислоты (реак-
ция восстановительного аминирования).
для систем защиты клетки от свободно-радикального окисления (анти-оксидантная защита).

2. Образование рибозо-5-фосфата, необходимого для синтеза нуклеиновых ки-
слот.
Наиболее активно реакции ПФП идут в цитозоле клеток печени, жировой ткани, эритроцитах, коре надпочечников, молочной железе при лактации, менее интенсив-
но в скелетных мышцах.
Пентозофосфатный путь включает два этапа – окислительный и неокислитель-ный.
На первом, окислительном, этапе глюкозо-6-фосфат в трех реакциях превра-
щается в рибулозо-5-фосфат, реакции сопровождаются восстановлением двух мо-лекул НАДФ до НАДФН.
Второй этап – этап структурных перестроек, благодаря которым пентозы воз-
вращаются в фонд гексоз. В этих реакциях рибулозо-5-фосфат может изомеризо-ваться до рибозо-5-фосфата и ксилулозо-5-фосфата. Далее под влиянием фермен-
тов транскетолазы и трансальдолазы происходят структурные перестройки с обра-
зованием иных моносахаридов. При реализации всех реакций второго этапа пентозы превращаются во фруктозо-6-фосфат и глицеральдегидфосфат. Из глицеральдегид-
3-фосфата при необходиости могут образоваться гексозы.
Связь пентозофосфатного пути и гликолиза
Судьба полученных фруктозо-6-фосфата и глицеральдегидфосфата различна
в зависимости от ситуации и потребностей клетки. Поэтому метаболизм глюкозо-6-фосфата может идти по 4 различным механизмам.
Механизм 1. Потребность в НАДФН и рибозо-5-фосфате сбалансирована
(например, при синтезе дезоксирибонуклеотидов). При таких условиях реак-
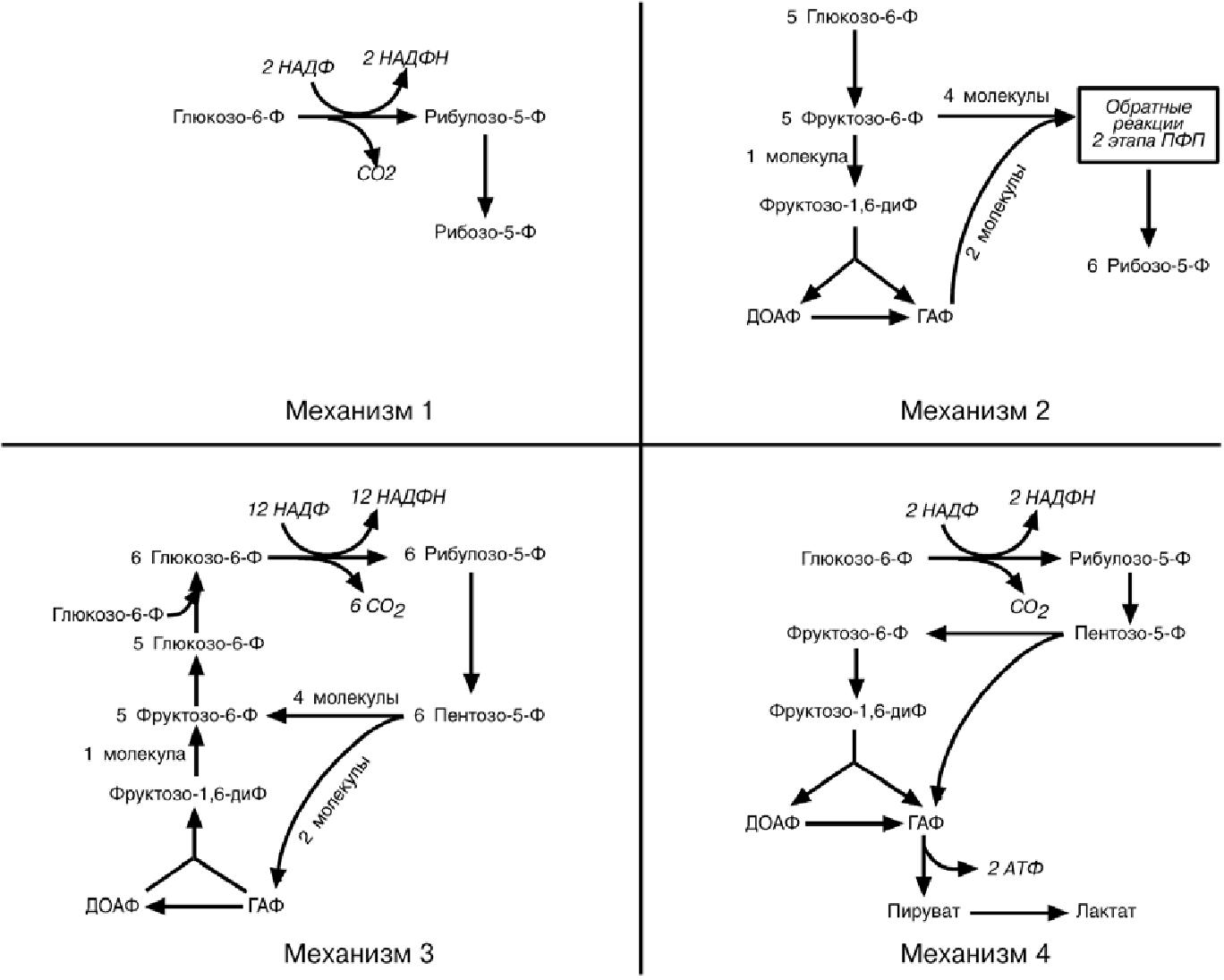
ции идут обычным порядком – образуется две молекулы NADPH и одна моле-
кула рибозо-5-фосфата из одной молекулы глюкозо-6-фосфата по окисли-тельной ветви пентозофосфатного пути.
Механизм 2. Потребность в рибозо-5-фосфате значительно превышает потребность в НАДФН (например, синтез РНК) Большая часть глюкозо-6-фосфата превращается во фруктозо-6-фосфат и глицеральдегид-3-фосфат
по гликолитическому пути. Затем две молекулы фруктозо-6-фосфата и одна молекула глицеральдегид-3-фосфата под действием трансальдолазы и
транскетолазы рекомбинируют в три молекулы рибозо-5-фосфата путем обращения реакции 2 этапа пентозофосфатного пути.
Механизм 3. Потребность в НАДФН значительно превышает потреб-
ность в рибозо-5-фосфате (например, биосинтез холестерола, жирных ки-слот). В этой ситуации по окислительным реакциям пентозофосфатного
пути образуются НАДФН и рибулозо-5-фосфат. Далее, под действием транс-кетолазы и трансальдолазы, рибулозо-5-фосфат превращается в пентозо-5-фосфаты, во фруктозо-6-фосфат и глицеральдегид-3-фосфат. В заключение
происходит ресинтез глюкозо-6-фосфата из фруктозо-6-фосфата и глице-ральдегидфосфата по пути глюконеогенеза. Подключение новых молекул
глюкозо-6-фосфата позволяет поддерживать стехиометрию процесса.
Механизм 4. Потребность в НАДФН значительно превышает потреб-ность в рибозо-5-фосфате и необходима энергия (например. антиокси-
дантная защита в эритроците). Глюкозо-6- фосфат превращается в рибозо-5-фосфат и далее во фруктозо-6- фосфат и глицеральдегид-3-фосфат, ко-торые ( в отличие от механизма 3) вступают на гликолитический путь обме-на, а не подвергаются обратному превращению в глюкозо-6-фосфат. Образо-ванный пируват вступает в ЦТК. В результате происходит одновременное генерирование НАДФН и АТФ.