

Рассмотрите важнейшие физиологические свойства нервных центров, обеспечивающие процессы адаптации к изменениям внешних условий или внутренней среды организма.
Нервный центр - совокупность нервных клеток выполняющих определенную функцию. Например, дыхательный нц, центр голода в гипоталамусе. В узком смысле слова – представлен группой нейронов, без участия которых функция будет неосуществима, в широком – включает участки НС, которые могут быть далеко от основной группы нейронов.
Свойства нервных центров: исходящие из свойств синапсов
одностороннее проведение возбуждения
центральная задержка (сумма синаптических задержек)
суммация (пространственная и временная)
низкая лабильность
трансформация ритма раздражителя - центры способны как снижать, так и повышать ритмы возбуждений, поступающих от рецепторов.
высокая утомляемость
высокая чувствительность к химическим веществам
свойство облегчения, долговременной потенциации, пластичности
тонус нервных центров (некоторое базальное возбуждение, которым отличаются нервные клети в составе НЦ) зависит от гуморальныхи афферентных влияний
доминанта (обнаружил Ухтомский) – тот нервный центр, тонус которого наиболее высоко притягивает к себе другие возбуждения от других нервных центров. Например, если имеется жажда и голод, то жажда будет доминировать и наличие голода будет усиливать жажду.
реципрокность – активность одного центра подавляет активность другого. Центр сгибателей и центр разгибателей.
исходящие из характера межнейрональных связей
конвергенция - ряда возбуждений к одному нейрону. Бывает мультисенсорная (свет и звук), сенсорно-биологическая (свет и голод), мультибиологическая (голод и боль)
дивергенция – от одного нейрона возбуждение распространяется на несколько нейронов o иррациация – распространение возбуждения от одного участка НЦ ко многим.
м
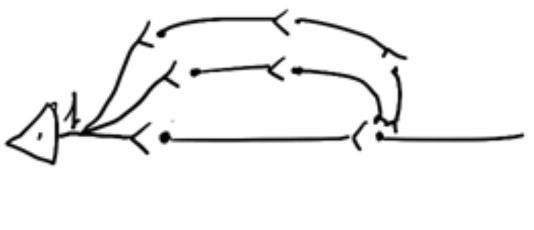 ультипликация
– к нейрону В не одновременно придут
потенциалы, потому что будет прибавляться
синаптическая задержка, а пути имеют
разную длину. Следовательно к нейрону
В придут 3 потенциала (умножение
возбуждения)
ультипликация
– к нейрону В не одновременно придут
потенциалы, потому что будет прибавляться
синаптическая задержка, а пути имеют
разную длину. Следовательно к нейрону
В придут 3 потенциала (умножение
возбуждения)
р
 еверберация
(есть теория, что так может осуществляться
коротко временная память)
еверберация
(есть теория, что так может осуществляться
коротко временная память)
П
 ринцип
окклюзии –
ринцип
окклюзии –
Дайте современное объяснение структуры связей нейронов и мембранных процессов, лежащих в основе известных видов центрального торможения. Объясните результаты опытов И.М.Сеченова (центральное торможение) и Гольца (сопряженное торможение) с современных позиций
Торможение – локальный активный процесс, который является следствием взаимодействия нескольких возбуждений.
Классификация по локализации (пресенаптическое и постсинаптическое) и по механизму торможения ( торможение, связанное с процессом гиперполяризации; пессимальное торможение, связанное с высокой частотой импульсации; торможение, связанное со стойкой деполяризацией – рефрактреным состоянием).
Клеточные виды торможения: возвратное торможение (физиологическая роль – препятствование перевозбуждению нейрона), латеральное (физиологическая роль – встречается в рецепторных полях и помогает различать/разделять/дискриминировать различные раздражители), реципрокное торможение – при возбуждении НЦ происходит реципрокное (сопряженное) торможение центра, отвечающего за противоположную функцию.
Сеченовское торможение. Наличие процесса торможения в ЦНС впервые было показано Сеченовым в 1862 г. в экспериментах на лягушке. Выполняли разрез головного мозга лягушки на уровне зрительных бугров и измеряли время рефлекса отдергивания задней лапы при погружении ее в раствор серной кислоты (рефлекс Тюрка). При наложении на разрез зрительных бугров кристаллика поваренной соли время рефлекса увеличивалось. Прекращение воздействия соли на зрительные бугры приводило к восстановлению исходного времени рефлекторной реакции. Рефлекс отдергивания лапки обусловлен возбуждением спинальных центров. Кристаллик соли, раздражая зрительные бугры, вызывает возбуждение, которое распространяется к спинальным центрам и тормозит их деятельность. И.М. Сеченов пришел к выводу, что торможение является следствием взаимодействия двух и более возбуждений на нейронах ЦНС. В этом случае одно возбуждение неизбежно становится тормозимым, а другое — тормозящим. Подавление одним возбуждением другого происходит как на уровне постсинаптических мембран (постсинаптическое торможение), так и за счет уменьшения эффективности действия возбуждающих синапсов на пресинаптическом уровне (пресинаптическое торможение).
Пресинаптическое торможение. Пресинаптическое торможение развивается в пресинаптической части синапса за счет воздействия на его мембрану аксо-аксональных синапсов. В результате как деполяризующего, так и гиперполяризующего воздействия происходит блокирование проведения импульсов возбуждения по пресинаптическим путям к постсинаптической нервной клетке.
Постсинаптическое торможение. Наибольшее распространение в ЦНС имеет механизм постсинаптического торможения, которое осуществляется специальными тормозными вставочными нервными клетками (например, клетки Реншоу в спинном мозге или клетки Пуркинье (грушевидные нейроны) в коре мозжечка). Особенность тормозных нервных клеток состоит в том, что в их синапсах имеются медиаторы, вызывающие на постсинаптической мембране нейрона ТПСП (тормозящие постсинаптические потенциалы), т.е. кратковременную гиперполяризацию. Например, для мотонейронов спинного мозга гиперполяризующим медиатором является аминокислота глицин, а для многих нейронов коры большого мозга таким медиатором служит гамма-аминомасляная кислота — ГАМК. Частным случаем постсинаптического является возвратное торможение.
Реципрокное торможение. Механизм постсинаптического торможения лежит в основе таких видов торможения, как реципрокное и латеральное. Реципрокное торможение является одним из физиологических механизмов координации деятельности нервных центров. Так, попеременно реципрокно тормозятся в продолговатом мозге центры вдоха и выдоха, прессорный и депрессорный сосудодвигательные центры. Реципрокное торможение проявляется на уровне спинного мозга при осуществлении строго координированных двигательных актов (ходьба, бег, чесание). На уровне сегментов спинного мозга возбуждение группы мотонейронов, вызывающих сокращение мышц-сгибателей, сопровождается реципрокным торможением другой группы мотонейронов, приводящих к расслаблению мышц-разгибателей. Латеральное торможение. Активность нейронов или рецепторов, расположенных рядом с возбужденными нейронами или рецепторами, прекращается. Механизм латерального торможения обеспечивает дискриминаторную способность анализаторов. Так, в слуховом анализаторе латеральное торможение обеспечивает различение частоты звуков, в зрительном анализаторе латеральное торможение резко увеличивает контрастность контуров воспринимаемого изображения, а в тактильном анализаторе способствует дифференцировке двух точек прикосновения.
При поступлении возбуждений к синапсам нервной клетки на постсинаптических мембранах могут возникать процессы гиперполяризации. Гиперполяризация приводит к возрастанию критического уровня деполяризации мембраны, следовательно, затрудняет возникновение возбуждения. Такие постсинаптические потенциалы получили название «тормозящие постсинаптические потенциалы» (ТПСП); они возникают в синапсах, где медиатор вызывает гиперполяризацию постсинаптической мембраны. Каждый нейрон синтезирует в своем теле и затем выделяет во всех своих синапсах один и тот же медиатор, поэтому нейроны и ацетилхолиновой передачей возбуждения называются холинергическими, с адреналиновой – адренергическими. К гиперполяризующим медиаторам относят ГАМК глицин. Эти медиаторы взаимодействуя с хеморецепторами постсинаптической мембраны, приводят к развитию ТПСП. Поступательное торможение обусловлено тем, что на пути следования возбуждения встречается тормозной нейрон.
Возвратное торможение осуществляется вставочными тормозными клетками (клетками Реншоу). Аксоны мотонейронов часто дают коллатерали (ответвления), оканчивающиеся на клетках Реншоу. Аксоны клеток Реншоу оканчиваются на теле или дендритах этого мотонейрона, образуя тормозные синапсы. Возбуждение, возникающее в мотонейроне, распространяется по прямому пути к скелетной мускулатуре, а также по коллатералям к тормозящему нейрону, которые посылают импульсы к мотонейронам и тормозят их. Чем сильнее возбуждение мотонейрона, тем сильнее возбуждаются клетки Реншоу и тем более интенсвно они оказывают свое тормозящее действие, что предохраняет нервные клетки от перевозбуждения.
Реципрокное (сопряженное) торможение. Это явление, открытое Ч. Шеррингтоном, обеспечивает согласованную работу мышц-антагонистов, например, сгибателей и разгибателей конечностей, т.е. движение конечностей. При сгибании ног в коленном суставе, развивается возбуждение в спинномозговом центре мышц-сгибателей и одновременно развивается торможение в нервном центре мышц-разгибателей.
Реципрокное торможение осуществляется при участии тормозящих вставочных нейронов спинного мозга. Ретикулярное торможение при раздражении ретикулярной формации кошка сгибает лапку.
Охарактеризуйте основные методы исследования ЦНС (электроэнцефалография, импульсная активность нейронов), объясните их использование для оценки функционального состояния человека (бодрствование, сон, мотивации, эмоции).
Методы исследования функций ЦНС делятся на две группы: 1) непосредственное изучение и 2) опосредованное (косвенное) изучение.
Методы непосредственного изучения функций ЦНС подразделяют на морфологические и функциональные. Морфологические методы. К морфологическим методам относятся макроанатомическое и микроскопическое исследования строения мозга. Этот принцип лежит в основе метода генетического картирования мозга, позволяющего выявлять функции генов в метаболизме нейронов. К морфологическим методам относят и метод меченых атомов. Сущность его заключается в том, что вводимые в организм радиоактивные вещества интенсивнее проникают в те нервные клетки мозга, которые в данный момент наиболее функционально активны.
Функциональные методы: разрушение и раздражение структур ЦНС, стереотаксический метод, электрофизиологические методы.
Метод разрушения. Разрушение структур мозга является довольно грубым методом исследования, поскольку повреждаются обширные участки мозговой ткани. Так, повреждение в области продолговатого мозга приводит к нарушению дыхания, глотания, деятельности сердца и изменению тонуса сосудов. В клинике для диагностики повреждений мозга различного происхождения (опухоли, инсульт и др.) у человека используют методы компьютерной рентгенотомографии, эхоэнцефалографии, ядерного магнитного резонанса.
Метод раздражения структур мозга позволяет установить пути распространения возбуждения от места раздражения к органу или ткани, функция которых при этом изменяется. В качестве раздражающего фактора чаще всего применяют электрический ток. В эксперименте на животных применяют метод самораздражения различных участков мозга: животное получает возможность посылать раздражение в мозг, замыкая цепь электрического тока и прекращать раздражение, размыкая цепь.
Стереотаксический метод введения электродов.
Стереотаксические атласы, которые имеют три координатных значения для всех структур мозга, помещенного в пространство трех взаимно перпендикулярных плоскостей — горизонтальной, сагиттальной и фронтальной. Данный метод позволяет не только с высокой точностью вводить электроды в мозг с экспериментальной и диагностической целями, но и направленно воздействовать на отдельные структуры ультразвуком, лазерными или рентгеновскими лучами с лечебной целью, а также проводить нейрохирургические операции. Электрофизиологические методы исследования ЦНС включают анализ как пассивных, так и активных электрических свойств мозга.
Электроэнцефалография. Метод регистрации суммарной электрической активности мозга называется электроэнцефалографией, а кривая изменений биопотенциалов мозга — электроэнцефалограммой (ЭЭГ). ЭЭГ регистрируют с помощью электродов, располагаемых на поверхности головы человека. Используют два способа регистрации биопотенциалов: биполярный и монополярный. При биполярном способе регистрируют разность электрических потенциалов между двумя близко расположенными точками на поверхности головы. При монополярном способе регистрируют разность электрических потенциалов между любой точкой на поверхности головы и индифферентной точкой на голове, собственный потенциал которой близок к нулю. Такими точками являются мочки уха, кончик носа, а также поверхность щек. Основными показателями, характеризующими ЭЭГ, являются частота и амплитуда колебаний биопотенциалов, а также фаза и форма колебаний. По частоте и амплитуде колебаний различают несколько видов ритмов в ЭЭГ.
Бета 13-30 Гц, амплитуда 14-40 мкВ, эмоциональное возбуждение, умственная и физическая деятельность, при нанесении раздражения.
Альфа 8-13 Гц, амплитуда до 100 мкВ состояние умственного и физического покоя, с закрытыми глазами.
Тета 4-8 Гц, амплитуда от 40 до 300 мкВ, промежуточная фаза сна, эмоциональное напряжение.
Дельта 1 – 4 Гц, амплитуда до 300 мкВ, глубокий сон, наркоз, гипоксия.
Основным и наиболее характерным ритмом является альфа-ритм. В состоянии относительного покоя альфаритм наиболее выражен в затылочных, затылочно-височных и затылочно-теменных областях головного мозга. При кратковременном действии раздражителей, например света или звука, появляется бета-ритм. Бета- и гамма-ритмы отражают активированное состояние структур головного мозга, тета-ритм чаще связан с эмоциональным состоянием организма. Дельта-ритм указывает на снижение функционального уровня коры большого мозга, связанное, например, с состоянием легкого сна или утомлением. Локальное появление дельта-ритма в какой-либо области коры мозга указывает на наличие в ней патологического очага.
Исследование импульсной активности нейронов с помощью микроэлектродной техники.
Электроды для регистрации клеточной активности часто изготавливаются из металла . В ходе операции под наркозом электроды вживляются в исследуемые структуры мозга и фиксируются к черепу животного. При регистрации импульсной активности животное осуществляет целенаправленное поведение . Анализ активности нервных клеток позволяет выявить особенности импульсации, характерной для того или иного поведения или состояния животного.
Исследование импульсной активности нейронов при микроионофорезе к нейронам биологически активных веществ.
Микроионофорез БАВ используется для изучения химической чувствительности мембран нервных клеток головного мозга. Регистрация осуществляется через центральный канал многоствольного стеклянного микроэлектрода. Биологически активные вещества в заряженном состоянии выводятся из боковых каналов электродов положительным или отрицательным током. Животное при этом может находится в свободном состоянии и осуществлять целенаправленное поведение.