

Гормональные механизмы регуляции почечной функции
34.3.1. В регуляции образования мочи в ответ на осмотический и другие сигналы принимают участие:
а) антидиуретический гормон;
б) система ренин-ангиотензин-альдостерон;
в) система предсердных натрийуретических факторов (атриопептидная система).
34.3.2. Антидиуретический гормон (АДГ, вазопрессин). АДГ синтезируется преимущественно в гипоталамусе в виде белка-предшественника, накапливается в нервных окончаниях задней доли гипофиза, из которых гормон секретируется в кровоток.
Сигналом для секреции АДГ служит повышение осмотического давления крови. Это может иметь место при недостаточном потреблении воды, сильном потоотделении или после приёма большого количества соли. Клетками-мишенями для АДГ являются клетки почечных канальцев, гладкомышечные клетки сосудов, а также клетки печени.
Влияние АДГ на почки заключается в удержании воды в организме путём стимуляции её реабсорбции в дистальных канальцах и собирательных трубочках. Взаимодействие гормона с рецептором активирует аденилатциклазу и стимулирует образование цАМФ. Под действием цАМФ-зависимой протеинкиназы фосфорилируются белки мембраны, обращённой в просвет канальца. Это придаёт мембране способность транспортировать в клетки воду, свободную от ионов. Вода поступает по градиенту концентрации, т.к. канальцевая моча гипотонична по отношению к содержимому клетки.
После приёма большого количества воды осмотическое давление крови снижается и синтез АДГ прекращается. Стенки дистальных канальцев становятся непроницаемыми для воды, реабсорбция воды уменьшается и, как следствие, выводится большой объём гипотонической мочи.
Заболевание, обусловленное дефицитом АДГ, получило название несахарный диабет. Оно может развиваться при нейротропных вирусных инфекциях, черепно-мозговых травмах, опухолях гипоталамуса. Основным симптомом этого заболевания является резкое увеличение диуреза (дол 10 и более литров в сутки) при пониженной (1,001-1,005) относительной плотности мочи.
34.3.3. Ренин-ангиотензин-альдостерон. Поддержание стабильной концентрации ионов натрия в крови и объём циркулирующей крови регулируется системой ренин-ангиотензин-альдостерон, которая влияет также на реабсорбцию воды. Уменьшение объёма крови, вызванное потерей натрия, стимулирует группу клеток, расположенных в стенках приносящих артериол - юкстагломерулярный аппарат (ЮГА). В него входят специализированные рецепторные и секреторные клетки. Активация ЮГА приводит к высвобождению из его секреторных клеток протеолитического фермента ренина. Ренин высвобождается из клеток также в ответ на снижение артериального давления крови.
Ренин воздействует на ангиотензиноген (белок a2-глобулиновой фракции), и расщепляет его с образованием декапептида ангиотензина I. Затем другой протеолитический фермент отщепляет от ангиотензина I два концевых аминокислотных остатка с образованием ангиотензина II. Этот октапептид является одним из наиболее активных средств, способствующих сужению кровеносных сосудов, в том числе артериол. В результате увеличивается кровяное давление, снижаются как почечный кровоток, так и клубочковая фильтрация.
Кроме того, ангиотензин II стимулирует секрецию клетками коркового слоя надпочечников гормона альдостерона. Альдостерон - гормон прямого действия - оказывает эффект на дистальный извитой каналец нефрона. Этот гормон индуцирует в клетках-мишенях синтез:
а) белков, участвующих в транспорте Na+ через люминальную поверхность клеточной мембраны;
б) Na+,K+-АТФазы, встраивающейся в контрлюминальную мембрану и участвующей в транспорте Na+ из клеток канальцев в кровь;
в) ферментов митохондрий, например, цитратсинтазы;
г) ферментов, участвующих в образовании фосфолипидов мембран, что облегчает транспорт Na+ в клетки канальца.
Таким образом, альдостерон повышает скорость реабсорбции Na+ из почечных канальцев (за ионами Na+ пассивно следуют ионы Cl—) и в конечном счёте осмотическую реабсорбцию воды, стимулирует активный перенос К+ из плазмы крови в мочу.
34.3.4. Предсердные натрийуретические факторы. Мышечные клетки предсердий синтезируют и секретируют в кровь пептидные гормоны, регулирующие диурез, экскрецию электролитов с мочой и сосудистый тонус. Эти гормоны получили название атриопептидов (от слова atrium - предсердие).
Атриопептиды млекопитающих независимо от размера молекулы имеют общую характерную структуру. У всех этих пептидов дисульфидная связь между двумя остатками цистеина образует 17-членную кольцевую структуру. Эта кольцевая структура обязательна для проявления биологической активности: восстановление дисульфидной группы приводит к потере активных свойств. От остатков цистеина отходят две пептидные цепи, представляющие N- и С-концевые участки молекулы. Количеством аминокислотных остатков на этих участках и отличаются друг от друга атриопептиды.
94
Особенности метаболизма эритроцита.
32.8.1. Эритроциты - высокоспециализированные клетки, основной функцией которых является транспорт кислорода из лёгких в ткани. Продолжительность жизни эритроцитов составляет в среднем 120 суток; разрушение их происходит в клетках ретикуло-эндотелиальной системы. В отличие от большинства клеток организма, у эритроцита отсутствуют клеточное ядро, рибосомы и митохондрии.
32.8.2. Энергетический обмен. Основным энергетическим субстратом эритроцита является глюкоза, которая поступает из плазмы крови путём облегчённой диффузии. Около 90% используемой эритроцитом глюкозы подвергается гликолизу (анаэробному окислению) с образованием конечного продукта - молочной кислоты (лактата). Запомните функции, которые выполняет гликолиз в зрелых эритроцитах:
1) в реакциях гликолиза образуется АТФ путём субстратного фосфорилирования. Основное направление использования АТФ в эритроцитах - обеспечение работы Na+,K+-АТФазы. Этот фермент осуществляет транспорт ионов Nа+ из эритроцитов в плазму крови, препятствует накоплению Na+ в эритроцитах и способствует сохранению геометрической формы этих клеток крови (двояковогнутый диск).
2) в реакции дегидрирования глицеральдегид-3-фосфата в гликолизе образуется НАДН. Этот кофермент является кофактором фермента метгемоглобинредуктазы,участвующей в восстановлении метгемоглобина в гемоглобин по следующей схеме:
![]()
Эта реакция препятствует накоплению метгемоглобина в эритроцитах.
3) метаболит гликолиза 1,3-дифосфоглицерат способен при участии фермента дифосфоглицератмутазы в присутствии 3-фосфоглицерата превращаться в 2,3-дифосфоглицерат:
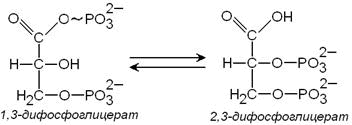
2,3-Дифосфоглицерат принимает участие в регуляции сродства гемоглобина к кислороду. Его содержание в эритроцитах повышается при гипоксии. Гидролиз 2,3-дифосфоглицерата катализирует фермент дифосфоглицератфосфатаза.
Приблизительно 10% глюкозы, потребляемой эритроцитом, используется в пентозофосфатном пути окисления. Реакции этого пути служат основным источником НАДФН для эритроцита. Данный кофермент необходим для перевода окисленного глутатиона (см. 30.8.3) в восстановленную форму. Дефицит ключевого фермента пентозофосфатного пути - глюкозо-6-фосфатдегидрогеназы - сопровождается уменьшением в эритроцитах отношения НАДФН/НАДФ+, увеличением содержания окисленной формы глутатиона и снижением резиcтентности клеток (гемолитическая анемия).
Механизмы обезвреживания активных форм кислорода в эритроцитах. Молекулярный кислород в определённых условиях может превращаться в активные формы, к которым относятся супероксидный анион О2-, пероксид водорода Н2О2, гидроксильный радикал ОН. и синглетный кислород 1О2. Эти формы кислорода обладают высокой реакционной способностью, могут оказывать повреждающее действие на белки и липиды биологических мембран, вызывать разрушение клеток. Чем выше содержание О2, тем больше образуется его активных форм. Поэтому эритроциты, постоянно взаимодействующие с кислородом, содержат эффективные антиоксидантные системы, способные обезвреживать активные метаболиты кислорода.
Важным компонентом антиоксидантных систем является трипептид глутатион, образующийся в эритроцитах в результате взаимодействия γ-глутамилцистеина и глицина:
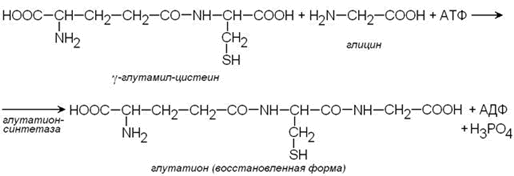
Восстановленная форма глутатиона (сокращённое обозначение Г-SH) участвует в реакциях обезвреживания пероксида водорода и органических пероксидов (R-O-OH). При этом образуются вода и окисленный глутатион (сокращённое обозначение Г-S-S-Г).

Превращение окисленного глутатиона в восстановленный катализирует фермент глутатионредуктаза. Источник водорода - НАДФН (из пентозофосфатного пути, см. 30.8.2):
![]()
В эритроцитах имеются также ферменты супероксиддисмутаза и каталаза, осуществляющие следующие превращения:

Антиоксидантные системы имеют для эритроцитов особое значение, так как в эритроцитах не происходит обновления белков путём синтеза.