

31-60
.docx31.
Биосинтез триацилглицеролов и глицерофосфолипидов происходит в цитоплазме клеток. Первые реакции этих метаболических путей совпадают и включают образование фосфатидной кислоты.
Предшественники фосфатидной кислоты - глицерол и жирные кислоты - вступают в реакцию в активной форме.
Образование активной формы глицерола (глицерол-3-фосфата может происходить двумя путями.
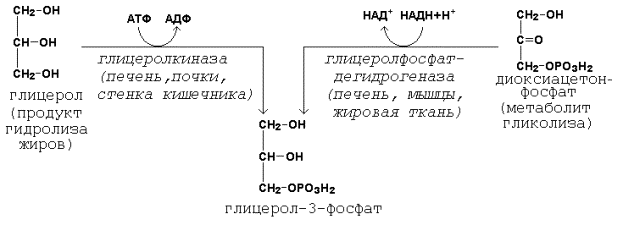
Пути образования глицерол-3-фосфата.
Активация жирных кислот идёт путём образования тиоэфиров коэнзима А – ацил-КоА .
Первая реакция ацилирования глицерол-3-фосфата осуществляется за счёт КоА-производного насыщенной жирной кислоты, например, пальмитиновой. Образуется сложноэфирная связь в 1-м положении.
Вторая реакция ацилирования идёт, как правило, за счёт КоА-тиоэфира ненасыщенной жирной кислоты, например, олеиновой.
В процессе синтеза триацилглицеролов происходит дефосфорилирование фосфатидной кислоты при помощи фосфатидатфосфатазы.
В дальнейшем 1,2-диацилглицерол ацилируется третьей молекулой ацил-КоА, которая может содержать остаток как ненасыщенной, так и насыщенной жирной кислоты.

Реакции биосинтеза триацилглицеролов.
В организме здорового человека процессы биосинтеза и катаболизма триацилглицеролов взаимно уравновешены.
В тех случаях, когда углеводы, жиры или белки потребляются в количествах, превышающих энергетические потребности организма, излишки калорий запасаются в виде триацилглицеролов. Источником ацетил-КоА для биосинтеза жирных кислот могут служить как углеводы, так и углеродные цепи аминокислот. Накопленный избыток жиров может быть израсходован для получения энергии, например, при голодании.
Скорость биосинтеза триацилглицеролов регулируется гормонами. Инсулин стимулирует поступление глюкозы в клетки и, следовательно, превращение её в жиры. Глюкагон и гормоны надпочечников тормозят этот процесс.
Глицерофосфолипиды являются производными фосфатидной кислоты и одного из аминоспиртов (например, этаноламина, холина или серина). Подобно другим предшественникам в биосинтезе липидов, аминоспирты, участвующие в синтезе фосфолипидов, вступают в реакцию в активной форме – в виде соединения с цитидиндифосфатом (ЦДФ). В качестве примера приводятся реакции активации этаноламина. Обратите внимание, что в реакциях активации аминоспиртов принимают участие два нуклеозидтрифосфата – АТФ и ЦТФ.
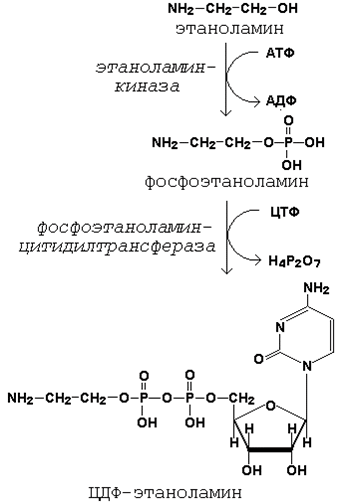
Реакции образования активной формы этаноламина.
Далее остаток фосфоэтаноламина с ЦДФ-этаноламина переносится на 1,2-диацилглицерол, образующийся в результате дефосфорилирования фосфатидной кислоты. Продуктом реакции является фосфатидилэтаноламин.

Реакции синтеза фосфатидилхолина.
Синтез фосфатидилхолина происходит путём трёхкратного метилирования фосфатидидэтаноламина. Донором метильных групп служит S-аденозилметионин. Фосфатидилхолин может образоваться также путём активации холина, подобно этаноламину.
Фосфатидилсерин образуется в реакции прямого взаимодействия фосфатидилэтаноламина и серина:
![]()
Липотропные факторы – вещества, способствующие синтезу фосфолипидов и препятствующие отложению триацилглицеролов в тканях.
Липотропный эффект этих соединений связан с тем, что общим предшественником триацилглицеролов и фосфолипидов является фосфатидная кислота. При недостатке липотропных факторов фосфатидная кислота используется преимущественно для синтеза триацилглицеролов. Они нерастворимы в воде и накапливаются в клетках, способствуя их жировому перерождению. Запомните, какие липотропные факторы можно использовать в качестве препаратов для его предупреждения:
1) холин – самое распространённое азотистое основание фосфолипидов;
2) метионин – незаменимая аминокислота, предшественник S-аденозилметионина, универсального донора метильных групп;
3) фолиевая кислота и витамин В12 – предшественники коферментов, участвующих в реакциях переноса СН3-групп.
При введении липотропных факторов снижается синтез триацилглицеролов и усиливается выведение жира из клеток при участии липопротеиновых комплексов
32.
Под термином «кетоновые тела» подразумевают следующие соединения: ацетоуксусная кислота (ацетоацетат), β-гидроксимасляная кислота (β-гидроксибутират), ацетон. Это – продукты неполного окисления жирных кислот. Синтез их происходит в митохондриях печени из ацетил-КоА.
(При β-окислении жирных кислот в митохондриях печени образуется ацетил-КоА, подвергающийся дальнейшему окислению в цикле Кребса. Если в клетках печени имеется дефицит оксалоацетата (например, при голодании, сахарном диабете), то происходит конденсация ацетильных групп с образованием кетоновых тел)
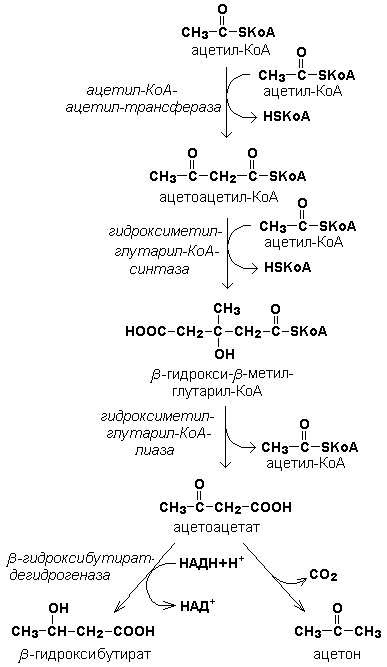
Реакции синтеза кетоновых тел.
Синтез кетоновых тел в организме усиливается при ускоренном катаболизме жирных кислот (голодание, сахарный диабет). В этих условиях в печени имеется дефицит оксалоацетата, образующегося преимущественно в реакциях углеводного обмена. Поэтому затрудняется взаимодействие ацетил-КоА с оксалоацетатом и все последующие реакции цикла трикарбоновых кислот Кребса.
Ацетоуксусная и β-гидроксимасляная кислоты, которые относятся к кетоновым телам, являются сильными кислотами. Поэтому накопление их в крови приводит к сдвигу рН в кислую сторону (метаболический ацидоз).
Кетоновые тела как источник энергии используют миокард, лёгкие, почки, скелетные мышцы, даже головной мозг (при длительном голодании). Утилизация кетоновых тел осуществляется следующим образом.
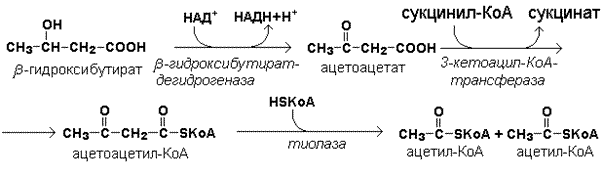
Реакции утилизации кетоновых тел.
Образующийся ацетил-КоА окисляется в цикле Кребса. Энергетический баланс окисления ацетоацетата до конечных продуктов составляет 23 молекулы АТФ, полного окисления β-гидроксибутирата - 26 молекул АТФ.
Печень не использует кетоновые тела в качестве энергетического материала.
При длительном голодании кетоновые тела становятся основным источником энергии для скелетных мышц, сердца и почек, а глюкоза потребляется мозгом и эритроцитами. Затем и мозг адаптируется к использованию ацетоуксусной кислоты. Если кетоновые тела накапливаются в крови в избытке (кетонемия), то они появляются в моче (кетонурия), а у лактирующих животных и в молоке (кетонолактия) - молоко становится горьким, непригодным для использования. Такое состояние называется кетозом. С потом, мочой, молоком, как правило, удаляется ацетон, который не утилизируется тканями. Именно ацетон создает своеобразный фруктовый запах животного или человека.
Гиперкетонемия представляет опасность для организма, так как приводит к ацидозу, сначала компенсированнному, с уменьшением щелочного резерва, а потом и к некомпенсированнному, со смещением рН. Накопление протонов в крови нарушает связывание кислорода гемоглобином и функцию других белков, в том числе ферментных. Возникают другие нарушения метаболизма, признаки сердечно-сосудистой недостаточности. У животных понижается или извращается аппетит, теряется масса, падает продуктивность, часто возникают аборты. При ацидозе кости теряют кальций, первыми признаками этого становятся рассасывание хвостовых позвонков и последних ребер, ломкость рогов. Гиперкетонемия может привести к кетоацидотической коме.
33.
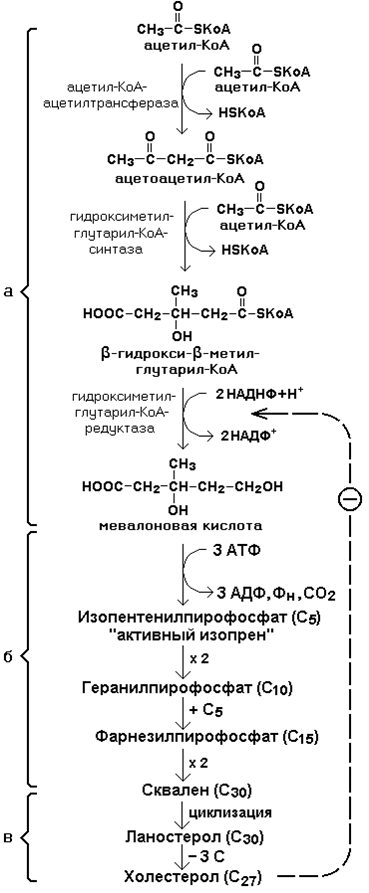
Исходным соединением для синтеза холестерола является ацетил-КоА. Ферменты, катализирующие реакции синтеза, содержатся в цитоплазме и эндоплазматическом ретикулуме многих клеток. Наиболее активно этот процесс происходит в печени. В организме человека в сутки синтезируется около одного грамма холестерола.
Биосинтез холестерола включает три основные стадии
Синтез холестерола.
На первой стадии образуется мевалоновая кислота.
На второй стадии мевалоновая кислота превращается в изопентенилпирофосфат («активный изопрен»), 6 молекул которого конденсируются в сквален.
На третьей стадии сквален превращается в холестерол.
Всего для синтеза 1 молекулы холестерола используется 18 молекул ацетил-КоА: для образования «активного изопрена» требуется 3 молекулы; в последующих реакциях конденсации участвуют 6 молекул «активного изопрена»; 3 × 6 = 18.
Скорость синтеза холестерола в организме регулируется по механизму отрицательной обратной связи (рисунок 21.5, пунктирная стрелка). Фермент β-гидрокси-β-метилглутарил-КоА-редуктаза катализирует лимитирующую реакцию биосинтеза холестерола. Холестерол является корепрессором синтеза данного ферментного белка, что приводит к снижению скорости катализируемой реакции. Поэтому при поступлении избытка холестерола с пищей синтез эндогенного холестерола прекращается.
Холестерол является компонентом биологических мембран, из него в организме образуются стероидные гормоны, витамин D3, желчные кислоты. Избыток холестерола превращается в печени в желчные кислоты, а также выделяется с желчью в кишечник и выводится с калом.
Нормальное содержание холестерола в сыворотке крови человека составляет 3,9 – 6,3 ммоль/л. Транспортной формой холестерола в крови являются липопротеины. Если нарушается соотношение между поступлением холестерола в организм и его выведением, то содержание холестерола в тканях и крови изменяется. Повышение концентрации холестерола в крови (гиперхолестеролемия) может приводить к развитию атеросклероза и желчно-каменной болезни.
34.
Окислению подвергаются все органические вещества. В ходе окислительных реакций органические молекулы разрушаются, а часть выделенной энергии запасается в виде АТФ.
Конечный продукт окислительных реакций – вода, но при этом образуются и так называемые активные формы кислорода – гидроксильный радикал, супероксидный анион, пероксид водорода. Они способны отнимать от органических молекул электроны, превращая их в активные радикалы и запуская таким образом цепные реакции повреждения молекул. В лейкоцитах, макрофагах такой механизм служит основой «респираторного взрыва», в ходе которого разрушаются бактерии и другие объекты фагоцитоза. Это полезная функция. Но в других клетках это приводит к саморазрушению органических молекул, в том числе и ДНК. Перекисное окисление липидов (ПОЛ), находящихся в клеточных мембранах, может привезти к гибели клеток. Наиболее подвержены действию активных форм кислорода непредельные жирные кислоты.
ПОЛ - цепные реакции, обеспечивающие расширенное воспроизводство свободных радикалов, частиц, имеющих неспаренный электрон, которые инициируют дальнейшее распространение перекисного окисления.
Стадии перекисного окисления липидов
1) Инициация: образование свободного радикала (L•)
Инициирует реакцию чаще всего гидроксильный радикал, отнимающий водород от СН2-групп полиеновой кислоты, что приводит к образованию липидного радикала.
2) Развитие цепи:
L • + О2 → LOO •
LOO• + LH → LOOM + LR•
Развитие цепи происходит при присоединении О2, в результате чего образуется липопе-роксирадикал LOO• или пероксид липида LOOH.
ПОЛ представляет собой свободнорадикальные цепные реакции, т.е. каждый образовавшийся радикал инициирует образование нескольких других.
3) Разрушение структуры липидов
Конечные продукты перекисного окисления полиеновых кислот - малоновый диальдегид и гидропероксид кислоты.
4) Обрыв цепи - взаимодействие радикалов между собой:
LOO• + L• → LOOH + LH
L• + vit E → LH + vit E•
vit E• + L• → LH + vit Еокисл.
Развитие цепи может останавливаться при взаимодействии свободных радикалов между собой или при взаимодействии с различными антиоксидантами, например, витамином Е, который отдаёт электроны, превращаясь при этом в стабильную окисленную форму.
Активные формы кислорода повреждают структуру ДНК, белков и различные мембранные структуры клеток. В результате появления в гидрофобном слое мембран гидрофильных зон за счёт образования гидропероксидов жирных кислот в клетки могут проникать вода, ионы натрия, кальция, что приводит к набуханию клеток, органелл и их разрушению. Активация перекисного окисления характерна для многих заболеваний: дистрофии мышц (болезнь Дюшенна), болезни Паркинсона, при которых ПОЛ разрушает нервные клетки в стволовой части мозга, при атеросклерозе, развитии опухолей. Перекисное окисление активируется также в тканях, подвергшихся сначала ишемии, а затем реоксигенации, что происходит, например, при спазме коронарных артерий и последующем их расширении.
ПОЛ разрушает клетки при атеросклерозе, развитии опухолей, нервные клетки, в которых много липидов. В организме имеются системы защиты клеток от активных форм кислорода: ферменты и витамины, обладающие антиоксидантным действием. Фермент супероксиддисмутаза (СОД) превращает супероксидные анионы в пероксид водорода. Фермент каталаза расщепляет пероксид водорода, который сам числится в повреждающих факторах. Фермент глутатионпероксидаза разрушает и пероксид водорода, и гидропероксиды липидов, предохраняя мембраны от повреждения. Коферментом глутатионпероксидазы является селен, поэтому его, как и витамины Е, С и β-каротины относят к факторам антиоксидантной защиты.
Арахидоновая кислота- эта 20-углеродная кислота содержит четыре двойные связи. Она является главным предшественником биологически активных соединений, объединяемых термином «простагландины». При окислении простагландинов в организме образуются два других биологических активных соединения — тромбоксан (в кровяных пластинках — тромбоцитах) и простациклин (в сосудистой стенке). Эти соединения влияют на агрегацию (образование конгломератов) и адгезию (прилипание к внутренней оболочке — эндотелию — сосудов) тромбоцитов, причем по-разному: тромбоксан ускоряет агрегацию и адгезию, а проста-циклин тормозит эти процессы. Поэтому соотношение тромбоксана и простациклина во многом определяет условия образования тромбов на поверхности эндотелия.
По строению фосфолипиды близки к триглицеридам, но отличаются от последних тем, что в их молекуле одна из трех частиц жирной кислоты заменена фосфорной кислотой (отсюда и название «фосфолипиды»), связанной с азотистым основанием. Фосфолипиды содержатся не только в крови, но и в мембранах всех клеток.
В клеточной мембране фосфолипиды вместе с холестерином выполняют роль «несущих конструкций» и ответственны за физическое состояние мембраны, точнее, за ее вязкость. От вязкости мембраны зависит активность мембранных ферментов и скорость переноса веществ через мембрану, другими словами, зависит функциональная активность клетки в целом.
Витамин Е (токоферол) - жирорастворимый витамин, антиоксидант, смесь из 4 токоферолов (от латинского tocos phero "потомство несущий") и 4 токотриенолов. Этот витамин был открыт в 1922 г. и получил пятый номер по порядку в алфавитной классификации, поэтому он имеет титул витамина Е. При дефиците его в рационе лабораторных крыс, у самок во время беременности погибал плод, а у самцов наступала тестикулярная атрофия.
Витамин Е существует, по крайней мере, в восьми различных конфигурациях, причем все они обладают той же активностью, что и "подлинная" молекула (d-альфа-токоферол), наиболее частая и активная форма. Она остается стандартом, с которым мы сравниваем активность других форм; активность их действия мы выражаем в эквивалентах d-альфа-токоферола.
Наибольшей витаминной активностью обладают α-β-γ-токоферолы (соотношение их эффективности составляет 100:40:8), а наибольшие антиоксидантные свойства присущи δ-токоферолу.
Недостаток витамина Е приводит к нарушению обмена веществ, снижению способности крови к передаче кислорода, развитию местного кислородного голодания. При недостаточном поступлении витамина Е наблюдается усиленное разрушение эритроцитов, связанное с нарушением стабильности их мембран. Витамин Е также улучшает циркуляцию крови, необходим для регенерации тканей, полезен при предменструальном синдроме и лечении фиброзных заболеваний груди. Он обеспечивает нормальную свертываемость крови и заживление; снижает возможность образования шрамов от некоторых ран; снижает кровяное давление; способствует предупреждению катаракт; улучшает атлетические достижения; снимает судороги ног; поддерживает здоровье нервов и мускулов; укрепляя стенки капилляров; предотвращает анемию.
В качестве антиоксиданта витамин Е защищает клетки от повреждения, замедляя окисление липидов (жиров) и формирование свободных радикалов. Он защищает другие растворимые жирами витамины от разрушения кислородом, способствует усвоению витамина А и защищает его от кислорода. Витамин Е замедляет старение, может предотвращать появление старческой пигментации.
Витамин Е участвует также в формировании коллагеновых и эластичных волокон межклеточного вещества. Токоферол предотвращает повышенную свертываемость крови, благоприятно влияет на периферическое кровообращение, участвует в биосинтезе гема и белков, пролиферации клеток, образовании гонадотропинов, развитии плаценты.
35.
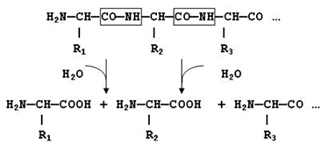
Переваривание белков, то есть расщепление их до отдельных аминокислот, начинается в желудке и заканчивается в тонком кишечнике. Переваривание происходит под действием желудочного, панкреатического и кишечного соков, которые содержат протеолитические ферменты (протеазы или пептидазы). Протеолитические ферменты относятся к классу гидролаз. Они катализируют гидролиз пептидных связей СО—NН белковой молекулы:
Гидролиз пептидных связей.
Все протеолитические ферменты можно разделить на две группы:
экзопептидазы – катализируют разрыв концевой пептидной связи с освобождением N- или С-концевой аминокислоты;
эндопептидазы – гидролизуют пептидные связи внутри полипептидной цепи, продуктами реакции являются пептиды с меньшей молекулярной массой.
Большинство протеолитических ферментов, участвующих в переваривании белков и пептидов, синтезируются и выделяются в полость пищеварительного тракта в виде неактивных предшественников – проферментов (зимогенов). Поэтому не происходит переваривания белков клеток, вырабатывающих проферменты. Активация проферментов осуществляется в просвете желудочно-кишечного тракта путём частичного протеолиза – отщепления части пептидной цепи зимогена.
Характеристика важнейших протеолитических ферментов
Фермент-Профермент-Источник-Активирующий фактор-Место действия-Оптимум рН-Специфичность действия
Пепсин-Пепсиноген-Желудочный сок-Соляная кислота, аутокатализ-Желудок-1,5 – 2,5-Эндопептидаза; разрыв связей: лей-глу; Х-фен; Х-тир
Трипсин-Трипсиноген-Панкреатический сок-Энтеропептидаза, аутокатализ-Тонкий кишечник-7,5 – 8,5-Эндопептидаза; разрыв связей: арг-Х; лиз-Х
Химотрипсин-Химотрипсиноген-Панкреатический сок-Трипсин-Тонкий кишечник-7,5 – 8,5-Эндопептидаза; разрыв связей: три-Х; фен-Х; тир-Х
Карбоксипептидаза-Прокарбоксипептидаза-Панкреатический сок-Трипсин-Тонкий кишечник-7,5 – 8,5-Экзопептидаза; отщепление С-концевых аминокислот
Аминопептидаза-Кишечный сок-Слизистая тонкого кишечника-7,5 – 8,5-Экзопептидаза; отщепление N-концевых аминокислот
(Х – любая аминокислота.)
Основная масса аминокислот, образовавшихся в пищеварительном тракте в результате переваривания белков, всасывается в кровь и пополняет аминокислотный фонд организма. Определённое количество невсосавшихся аминокислот подвергается гниению в толстом кишечнике.
Гниение – превращения аминокислот, вызванные деятельностью микроорганизмов в толстом кишечнике. Усилению процессов гниения аминокислот могут способствовать:
-избыточное поступление белков с пищей;
-врождённые и приобретённые нарушения процесса всасывания аминокислот в кишечнике;
-снижение моторной функции кишечника.
В результате гниения аминокислот образуются различные вещества, многие из которых являются токсичными для организма.
Продукты гниения аминокислот в кишечнике.(Аминокислоты-Продукты гниения)
Тирозин-Крезол, Фенол
Триптофан-Скатол, Индол
Цистеин, Метионин-Метилмеркаптан, Сероводород
Лизин-Кадаверин
Орнитин-Путресцин
Продукты гниения аминокислот являются ксенобиотиками – веществами, чужеродными для организма человека и должны быть обезврежены (инактивированы).
Обезвреживание продуктов гниения аминокислот происходит в клетках печени после поступления веществ из кишечника с кровью воротной вены. Продукты обезвреживания хорошо растворяются в воде и поэтому легко выводятся из организма. Процесс обезвреживания включает, как правило, две фазы (стадии): фазу модификации и фазу конъюгации.
36.
Декарбоксилирование аминокислот – отщепление карбоксильной группы от аминокислоты с образованием СО2. Продуктами реакций декарбоксилирования аминокислот являются биогенные амины, участвующие в регуляции обмена веществ и физиологических процессов в организме.
Биогенные амины и их предшественники(Аминокислота-Биогенный амин)
Гистидин-Гистамин
Глутамат-γ-аминомасляная кислота (ГАМК)
Тирозин-Дофамин
Триптофан-Триптамин, Серотонин
Цистеин-Тиоэтиламин, Таурин
Реакции декарбоксилирования аминокислот и их производных катализируют декарбоксилазы аминокислот. Кофермент – пиридоксальфосфат (производное витамина В6). Реакции являются необратимыми.
Примеры реакций декарбоксилирования. Некоторые аминокислоты непосредственно подвергаются декарбоксилированию. Реакция декарбоксилирования гистидина:

Гистамин обладает мощным сосудорасширяющим действием, особенно капилляров в очаге воспаления; стимулирует желудочную секрецию как пепсина, так и соляной кислоты, и используется для исследования секреторной функции желудка.
Реакция декарбоксилирования глутамата:

ГАМК – тормозный медиатор в центральной нервной системе.
Ряд аминокислот подвергается декарбоксилированию после предварительного окисления. Продукт гидроксилирования триптофана превращается в серотонин:

Серотонин образуется главным образом в клетках центральной нервной системы, обладает сосудосуживающим действием. Участвует в регуляции артериального давления, температуры тела, дыхания, почечной фильтрации.
Продукт гидроксилирования тирозина переходит в дофамин.Дофамин служит предшественником катехоламинов; является медиатором ингибирующего типа в центральной нервной системе.
Таурин образуется главным образом в печени; участвует в синтезе парных желчных кислот (таурохолевой кислоты).
Катаболизм биогенных аминов. В органах и тканях существуют специальные механизмы, предупреждающие накопление биогенных аминов. Основной путь инактивации биогенных аминов – окислительное дезаминирование с образованием аммиака – катализируется моно- и диаминооксидазами.
Моноаминооксидаза (МАО) - ФАД-содержащий фермент – осуществляет реакцию:
![]()
В клинике используются ингибиторы МАО (ниаламид, пиразидол) для лечения депрессивных состояний.
37.
Дезаминирование аминокислот – отщепление аминогруппы от аминокислоты с образованием аммиака (NН3). В тканях человека преобладает окислительное дезаминирование, то есть сопряжённое с переносом водорода.
Большинство ферментов, участвующих в окислительном дезаминировании аминокислот, при физиологических значениях рН малоактивны. Поэтому основная роль в окислительном дезаминировании принадлежит глутаматдегидрогеназе, которая катализирует прямое окислительное дезаминирование глутамата. В качестве кофермента используются НАД+ или НАДФ+ (производные витамина РР). Реакция обратима.
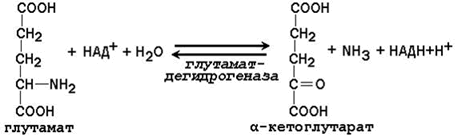
Глутаматдегидрогеназа – аллостерический фермент, его аллостерическими активаторами являются АДФ и ГДФ, аллостерическими ингибиторами – АТФ, ГТФ и НАДН.
Непрямое дезаминирование характерно для большинства аминокислот. Оно называется непрямым, потому что происходит в 2 этапа:
-на первом этапе аминокислота подвергается трансаминированию с образованием глутамата;
-на втором этапе происходит окислительное дезаминирование глутамата.

Схема непрямого дезаминирования аминокислот.
Участие аминотрансфераз в этом процессе позволяет собрать аминогруппы различных аминокислот в составе одной аминокислоты – глутамата, который затем подвергается окислению с образованием аммиака и α-кетоглутарата.
Трансаминирование аминокислот – перенос аминогруппы (NН2-) от аминокислоты на α-кетокислоту без промежуточного образования аммиака. Реакции трансаминирования катализируют ферменты – аминотрансферазы (или трансаминазы). Кофермент аминотрансфераз – пиридоксальфосфат (производное витамина В6). В реакции принимает участие альдегидная группа кофермента. Реакция легко обратима.
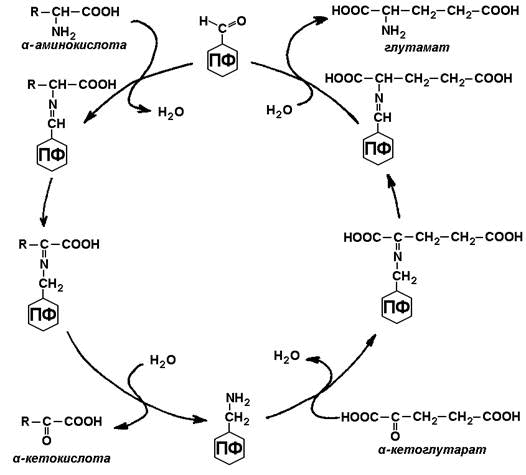
Механизм переноса аминогруппы с аминокислоты на α-кетокислоту в реакции трансаминирования.
Примеры реакций трансаминирования:
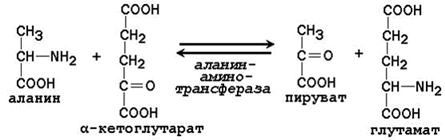

Роль реакций трансаминирования в организме:
участие в непрямом дезаминировании аминокислот;
путь синтеза заменимых аминокислот;
образующиеся в реакции α-кетокислоты могут включаться в общий путь катаболизма и глюконеогенез.
38.
Аммиак (NН3) – продукт обмена большинства соединений, содержащих амино- и амидогруппы. Главным путём образования аммиака служит окислительное дезаминирование.