

Особенности образования аммиака
-кетоглутаровая кислота. Считается, что при увеличении концентрации аммиака в крови значительная часть этой кислоты используется для связывания аммиака. В результате происходит “утечка” субстратов из цикла трикарбоновых кислот. Это, в свою очередь, нарушает процессы окисления и ухудшает энергообеспеченность нервных клеток.Аммиак образуется в мозге, главным образом, при участии аденилатдезаминазы (рис. 18.6). Атом азота аминокислоты через систему глутамат - аспартат попадает в аденилат (АМФ), который и дезаминируется. Аммиак оказывает токсическое действие на функции нейронов. Это связано с особенностями механизмов его обезвреживания в нервной ткани. Основное место в обезвреживании аммиака занимают реакции образования глютамина. В этом процессе принимают участие глутаматдегидрогеназа и глутаминсинтетаза. В качестве исходного субстрата для образования глутамина используется важный промежуточный продукт цикла трикарбоновых кислот - Нервная ткань характеризуется высоким содержанием РНК и довольно большой скоростью образования этих молекул. В ткани мозга содержится полный набор ферментов синтеза de novo пуриновых нуклеотидов, а синтез de novo пиримидиновых нуклеотидов невозможен из-за отсутствия карбомоилфосфатсинтетазы. Зато нуклеозиды легко проходят гемато-энцефалический барьер и могут повторно включаться в синтез нуклеотидов. Недостаток одного из ферментов, катализирующих повторное использование нуклеозидов, приводит к тяжелому нарушению функций мозга (синдром Леш-Нихана). ^
Особенности обмена липидов
Нервная ткань отличается высокой интенсивностью обмена липидов в период развития организма и относительной стабильностью обмена у взрослого. Как уже указывалось, скорость обновления липидов мозга довольно низкая. Длительное голодание несущественно влияет на липидный обмен нервной ткани. В молодом возрасте нервные клетки способны синтезировать холестерол, однако в последующем идет постепенное снижение активности гидроксиметилглутарилредуктазы, замедление и прекращение синтеза холестерола. Активное образование сложных липидов идет в период миелинизации. Врожденные нарушения обмена сложных липидов сопровождаются тяжелыми нарушениями функций мозга (см. главу "Обмен липидов"). ^
Нейромедиаторы - это вещества, которые характеризуются следующими признаками.
Накапливаются в пресинаптической структуре в достаточной концентрации.
Освобождаются при передаче импульса.
Вызывают после связывания с постсинаптической мембраной изменение скорости метаболических процессов и возникновение электрического импульса.
Имеют систему для инактивации или транпортную систему для удаления из синапса, обладающие к ним высоким сродством.
Таким образом, нейромедиаторы играют важную роль в функционировании нервной ткани, обеспечивая синаптическую передачу нервного импульса. Их синтез происходит в теле нейронов, а накопление - в особых везикулах, которые постепенно перемещаются с участием систем нейрофиламентов и нейротрубочек к кончикам аксонов.
Химическая классификация нейромедиаторов. Аминокислоты (и их производные).
К ним относят таурин, норадреналин, ДОФАминГАМК, глицин, ацетилхолин, гомоцистеин и некоторые другие (адреналин, серотонин, гистамин, серотонин).
Таурин.
Таурин образуется из аминокислоты цистеина. Сначала происходит окисление серы в SH-группе до остатка серной кислоты (процесс идет в несколько стадий), а затем происходит декарбоксилирование. Таурин - это необычная кислота, в которой нет карбоксильной группы, а имеется остаток серной кислоты.
Таурин принимает участие в проведении нервного импульса в процессе зрительного восприятия.
Ацетилхолин.
Для синтеза холина требуются аминокислоты серин, метионин. Этаноламин может быть использован и в готовом виде. Но, как правило, из крови в нервную ткань пступает уже готовый холин. Второй же предшественник этого нейромедиатора - Ацетил-КоА, синтезируется в нервных окончаниях.
Продукт этой реакции ацетилхолин участвует в синаптической передаче нервного импульса. Он накапливается в синаптических пузырьках, образуя комплексы с отрицательно заряженным белком везикулином. Передача возбуждения с одной клетки на другую осуществляется с помощью специального синаптического механизма.
Синапс - это функциональный контакт специализированных участков плазматических мембран двух возбудимых клеток. Синапс состоит из пресинаптической мембраны, синаптической щели и постинаптической мембраны. Мембраны клеток в месте контакта имеют утолщения в виде бляшек - нервных окончаний. Нервный импульс, достигший нервного окончания, не в состоянии преодолеть возникшее перед ним препятствие - синаптическую щель. После этого электрический сигнал преобразуется в химический. Пресинаптическая мембрана содержит специальные канальные белки, подобные белкам, формирующим натриевый канал в мембране аксона. Они тоже реагируют на мембранный потенциал, изменяя свою конформацию и формируют канал. В результате ионы Са2+ проходят через пресинаптическую мембрану по градиенту концентраций в нервное окончание. Градиент концентраций Са2+ создается работой Са2+-зависимой.
АТФазы - кальциевым насосом. Повышение концентрации Са2+ внутри нервного окончания вызывает слияние 200-300 имеющихся там везикул, заполненных ацетилхолином, с плазматической мембраной. Далее ацетилхолин секретируется в синаптическую щель путем экзоцитоза, и присоединяется к рецепторным белкам, расположенным на поверхности постсинаптической мембраны.
Ацетилхолиновый рецептор представляет собой трансмембранный олигомерный гликопротеиновый комплекс, состоящий из 6 субъединиц: 2-, 2-бета, 1-гамма и 1-дельта. Плотность расположения белков-рецепторов в постсинаптической мембране очень велика - около 20000 молекул на 1 мкм2. Пространственная структура рецептора строго сооответствует конформации медиатора.
При взаимодействии с ацетилхолином белок-рецептор так изменяет свою конформацию, что внутри него формируется натриевый канал. Катионная селективость канала обеспечивается тем, что ворота канала сформированы отрицательно заряженными аминокислотами. Таким образом повышается проницаемость постсинаптической мембраны для натрия и возникает новый импульс (или сокращение мышечного волокна). Деполяризация постсинаптической мембраны выеывает диссоциацию комплекса "ацетилхолин-белок-рецептор" и ацетилхолин освобождается в синаптическую щель. Как только ацетилхолин оказывается в синаптической щели, он за 40 мкс подвергается быстрому гидролизу под действием фермента ацетилхолинэстеразы.
Во время гидролиза ацетилхолина образуется промежуточный фермент-субстратный комплекс, в котором ацетилхолин связан с активным центром фермента через серин.
Необратимое ингибирование холинэстеразы вызывает смерть. Ингибиторами холинэстеразы являются фосфорорганические соединения (хлорофос, дихлофос, табун, зарин, зоман, бинарные яды). Эти вещества связываются ковалентно с серином в активном центре фермента. Некоторые из них синтезированы в качестве инсектицидов, а некоторые - в качестве боевых отравляющих веществ (нервно-паралитические яды). Смерть наступает в результате остановки дыхания.
Обратимые ингибиторы холинэстеразы используются как лечебные препараты. Например, при лечении глаукомы и атонии кишечника.
Катехоламины: норадреналин и дофамин. Адренэргические синапсы встречаются в постганглионарных волокнах, в волокнах симпатической нервной системы, в различных отделах головного мозга. Катехоламины в нервной ткани синтезируются по общему механизму из тирозина. Ключевой фермент синтеза - тирозингидроксилаза, ингибируемая конечными продуктами.
Норадреналин - медиатор в постганглионарных волокнах симпатической и в различных отделах ЦНС.
Дофамин- медиатор проводящих путей, тела нейронов которого расположены в отделе мозга, который отвечает за контроль произвольных движений. Поэтому при нарушении дофаминэргической передачи возникает заболевание паркинсонизм.
Катехоламины, как и ацетилхолин, накапливаются в синаптических пузырьках и тоже выделяется в синаптическую щель при поступлении нервного импульса. Но регуляция в адренэргическом рецепторе происходит иначе. В пресинаптической мембране здесь имеется специальный регуляторный белок - -ахромогранин (Мм = 77 кДа), который в ответ на повышение концентрации медиатора в синаптической щели связывает уже выделившийся медиатор и прекращает его дальнейший экзоцитоз. Фермента, разрушающего медиатор, в адренэргических синапсах нет. После передачи импульса молекулы медиатора перекачивается специальной транспортной системой путем активного транспорта с участием АТФ обратно через пресинаптическую мембрану и включается вновь в везикулы. В пресинаптическом нервном окончании излишек медиатора может быть инактивирован моноаминоксидазой, а также катехоламин-О-метилтрансферазой путем метилирования по оксигруппе. Кокаин тормозит активный транспорт катехоламинов.
Передача сигнала в адренэргических синапсах протекает по механизму, известному Вам из лекций по теме "Биохимия гормонов" с участием аденилатциклазной системы. Связывание медиатора с постсинаптическим рецептором почти мгновенно вызывает повышение концентрации ц-АМФ, что приводит к быстрому фосфорилированию белков постсинаптической мембраны. В результате изменяется генерация нервных импульсов постсинаптической мембраной (тормозится). В некторых случаях непосредственной причиной этого является повышение проницаемости постсинаптической мембраны для калия, либо снижением проводимости для натрия (эти события приводят к гиперполяризации).
ГАМК - тормозной медиатор. Повышает проницаемость постсинаптических мембран для ионов калия. Это ведет к изменению мембранного потенциала.
Глицин - тормозной медиатор, по вызываемым эффектам подобен гамк.
Пептиды.
Имеют в своем составе от трех до нескольких десятков аминокислотных остатков. Функциониуют только в высших отделах нервной системы.
Эти пептиды, как и катехоламины, выполняют функцию не только нейромедиаторов, но и гормонов. Передают информацию от клетки к клетке по системе циркуляции.
99
Белки плазмы крови.
Плазма крови содержит сложную многокомпонентную (более 100) смесь белков, различающихся по происхождению и функциям. Большинство белков плазмы синтезируется в печени. Иммуноглобулины и ряд других защитных белков иммунокомпетентными клетками.
Содержание общего белка в сыворотке крови здорового человека составляет 65 - 85 г/л (в плазме крови этот показатель на 2 – 4 г/л выше за счёт фибриногена).
32.2.1. Белковые фракции. При помощи высаливания белков плазмы можно выделить альбуминовую и глобулиновую фракции. В норме соотношение этих фракций составляет 1,5 – 2,5. Использование метода электрофореза на бумаге позволяет выявить 5 белковых фракций (в порядке убывания скорости миграции): альбумины, α1-, α2-, β- и γ-глобулины. При использовании более тонких методов фракционирования в каждой фракции, кроме альбуминовой, можно выделить целый ряд белков (содержание и состав белковых фракций сыворотки крови см. рисунок 1).
 Рисунок
1. Электрофореграмма
белков сыворотки крови и состав белковых
фракций.
Рисунок
1. Электрофореграмма
белков сыворотки крови и состав белковых
фракций.
Альбумины – белки с молекулярной массой около 70000 Да. Благодаря гидрофильности и высокому содержанию в плазме играют важную роль в поддержании коллоидно-осмотического (онкотического) давления крови и регуляции обмена жидкостей между кровью и тканями. Выполняют транспортную функцию: осуществляют перенос свободных жирных кислот, желчных пигментов, стероидных гормонов, ионов Са2+, многих лекарств. Альбумины также служат богатым и быстро реализуемым резервом аминокислот.
α1-Глобулины:
Кислый α1-гликопротеин (орозомукоид) – содержит до 40% углеводов, изоэлектрическая точка его находится в кислой среде (2,7). Функция этого белка до конца не установлена; известно, что на ранних стадиях воспалительного процесса орозомукоид способствует образованию коллагеновых волокон в очаге воспаления (Я.Мусил, 1985).
α1-Антитрипсин – ингибитор ряда протеаз (трипсина, химотрипсина, калликреина, плазмина). Врождённое снижение содержания α1-антитрипсина в крови может быть фактором предрасположенности к бронхо-лёгочным заболеваниям, так как эластические волокна лёгочной ткани особенно чувствительны к действию протеолитических ферментов.
Ретинолсвязывающий белок осуществляет транспорт жирорастворимого витамина А.
Тироксинсвязывающий белок – связывает и транспортирует иодсодержащие гормоны щитовидной железы.
Транскортин – связывает и транспортирует глюкокортикоидные го рмоны (кортизол, кортикостерон).
α2-Глобулины:
Гаптоглобины (25% α2-глобулинов) – образуют стабильный комплекс с гемоглобином, появляющимся в плазме в результате внутрисосудистого гемолиза эритроцитов. Комплексы гаптоглобин-гемоглобин поглощаются клетками РЭС, где гем и белковые цепи подвергаются распаду, а железо повторно используется для синтеза гемоглобина. Тем самым предотвращается потеря железа организмом и повреждение почек гемоглобином.
Церулоплазмин – белок, содержащий ионы меди (одна молекула церулоплазмина содержит 6-8 ионов Cu2+), которые придают ему голубую окраску. Является транспортной формой ионов меди в организме. Обладает оксидазной активностью: окисляет Fe2+ в Fe3+, что обеспечивает связывание железа трансферрином. Способен окислять ароматическиеамины, участвует в обмене адреналина, норадреналина, серотонина.
β-Глобулины:
Трансферрин – главный белок β-глобулиновой фракции, участвует в связывании и транспорте трёхвалентного железа в различные ткани, особенно в кроветворные. Трансферрин регулирует содержание Fe3+ в крови, предотвращает избыточное накопление и потерю с мочой.
Гемопексин – связывает гем и предотвращает его потерю почками. Комплекс гем-гемопексин улавливается из крови печенью.
С-реактивный белок (С-РБ) – белок, способный преципитировать (в присутствии Са2+) С-полисахарид клеточной стенки пневмококка. Биологическая роль его определяется способностью активировать фагоцитоз и ингибировать процесс агрегации тромбоцитов. У здоровых людей концентрация С-РБ в плазме ничтожно мала и стандартными методами не определяется. При остром воспалительном процессе она увеличивается более чем в 20 раз, в этом случае С-РБ обнаруживается в крови. Исследование С-РБ имеет преимущество перед другими маркерами воспалительного процесса: определением СОЭ и подсчётом числа лейкоцитов. Данный показатель более чувствителен, его увеличение происходит раньше и после выздоровления быстрее возвращается к норме.
γ-Глобулины:
Иммуноглобулины (IgA, IgG, IgM, IgD, IgE) представляют собой антитела, вырабатываемые организмом в ответ на введение чужеродных веществ с антигенной активностью. Подробнее об этих белках см. 1.2.5.
32.2.2. Количественные и качественные изменения белкового состава плазмы крови. При различных патологических состояниях белковый состав плазмы крови может изменяться. Основными видами изменений являются:
Гиперпротеинемия - увеличение содержания общего белка плазмы. Причины: потеря большого количества воды (рвота, диарея, обширные ожоги), инфекционные заболевания (за счёт увеличения количества γ-глобулинов).
Гипопротеинемия - уменьшение содержания общего белка в плазме. Наблюдается при заболеваниях печени (вследствие нарушения синтеза белков), при заболеваниях почек (вследствие потери белков с мочой), при голодании (вследствие недостатка аминокислот для синтеза белков).
Диспротеинемия - изменение процентного соотношения белковых фракций при нормальном содержании общего белка в плазме крови, например, снижение содержания альбуминов и увеличение содержания одной или нескольких глобулиновых фракций при различных воспалительных заболеваниях.
Парапротеинемия - появление в плазме крови патологических иммуноглобулинов - парапротеинов, отличающихся от нормальных белков по физико-химическим свойствам и биологической активности. К таким белкам относятся, например, криоглобулины, образующие друг с другом преципитаты при температуре ниже 37° С. Парапротеины обнаруживаются в крови при макроглобулинемии Вальденстрема, при миеломной болезни (в последнем случае они могут преодолевать почечный барьер и обнаруживаться в моче как белки Бенс-Джонса). Парапротеинемия, как правило, сопровождается гиперпротеинемией.
. Белки острой фазы воспаления. Это белки, содержание которых увеличивается в плазме крови при остром воспалительном процессе. К ним относятся, например, следующие белки:
гаптоглобин;
церулоплазмин;
С-реактивный белок;
α1-антитрипсин;
фибриноген (компонент свёртывающей системы крови; см. 30.7.2).
Скорость синтеза этих белков увеличивается прежде всего за счёт снижения образования альбуминов, трансферрина и альбуминов (небольшая фракция белков плазмы, обладающая наибольшей подвижностью при диск-электрофорезе, и которой соответствует полоса на электрофореграмме перед альбуминами), концентрация которых при остром воспалении снижается.
Биологическая роль белков острой фазы: а) все эти белки являются ингибиторами ферментов, освобождаемых при разрушении клеток, и предупреждают вторичное повреждение тканей; б) эти белки обладают иммунодепрессорным действием (В.Л.Доценко, 1985).
32.2.5. Защитные белки плазмы крови. К белкам, выполняющим защитную функцию, относятся иммуноглобулины и интерфероны.
Иммуноглобулины (антитела) - группа белков, вырабатываемых в ответ на попадание в организм чужеродных структур (антигенов). Они синтезируются в лимфоузлах и селезёнке лимфоцитами В. Выделяют 5 классов иммуноглобулинов - IgA, IgG, IgM, IgD, IgE.
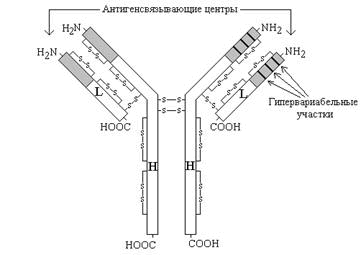 Рисунок
3. Схема
строения иммуноглобулинов (серым цветом
показана вариабельная область, не
закрашена - константная область).
Рисунок
3. Схема
строения иммуноглобулинов (серым цветом
показана вариабельная область, не
закрашена - константная область).
Молекулы иммуноглобулинов имеют единый план строения. Структурную единицу иммуноглобулина (мономер) образуют четыре полипептидные цепи, соединённые между собой дисульфидными связями: две тяжёлые (цепи Н) и две лёгкие (цепи L) (см. рисунок 3). IgG, IgD и IgЕ по своей структуре, как правило, являются мономерами, молекулы IgM построены из пяти мономеров, IgA состоят из двух и более структурных единиц, или являются мономерами.
Белковые цепи, входящие в состав иммуноглобулинов, можно условно разделить на специфические домены, или области, имеющие определённые структурные и функциональные особенности.
N-концевые участки как L-, так и Н-цепей называются вариабельной областью (V), так как их структура характеризуется существенными различиями у разных классов антител. Внутри вариабельного домена имеются 3 гипервариабельных участка, отличающихся наибольшим разнообразием аминокислотной последовательности. Именно вариабельная область антител ответственна за связывание антигенов по принципу комплементарности; первичная структура белковых цепей в этой области определяет специфичность антител.
С-концевые домены Н- и L-цепей обладают относительно постоянной первичной структурой в пределах каждого класса антител и называются константной областью (С). Константная область определяет свойства различных классов иммуноглобулинов, их распределение в организме, может принимать участие в запуске механизмов, вызывающих уничтожение антигенов.