

27. Меристеми. Типи меристем та галузі їх використання у біотехнології. Принципи складання поживних середовищ для культивування клітин.
Меристема (від грецьк. meristos – той, що здатний до поділу) – це група клітин, яка зберігає здатність до мітотичного поділу з утворенням дочірніх клітин. Ці клітини ростуть, створюючи постійну тканину, що складається з клітин, вже не здатних до поділу. Існує три типи меристем: апікальна, латеральна, інтеркалярна. Їх характеристику наведено у таблиці 6.
Клітини меристеми мають невеликий розмір, тонку оболонку і велике ядро. Вони щільно прилягають одна до одної без міжклітинного простору. Типова клітина апікальної меристеми – відносно дрібна кубоподібна клітина з тонкою целюлозною стінкою та густою цитоплазмою.
Типи меристем та їх функції
|
Тип меристеми |
Місце знаходження |
Роль |
езультат |
|
Апікальна |
У кінчиках коренів та пагонів |
Забезпечує первинний ріст, утворюючи первинне тіло рослини |
Збільшення у довжину
|
|
Латеральна (камбій) |
У старіших частинах рослини, розташовується паралельно довгій осі органа, наприклад короковий камбій (фелоген) та судинний камбій |
Забезпечує вторинний ріст. Судинний камбій дає початок вторинним провідним тканинам, у т.ч. деревині (вторинній ксилемі); короковий камбій утворює перидерму, яка заміняє епідерміс і містить корок |
Збільшення у товщину |
|
Інтеркалярна |
Між ділянками постійних тканин, наприклад у вузлах у багатьох однодольних – в основі листків у злаків |
Робить можливим ріст у проміжних ділянках (не на кінчиках). Це суттєво для тих рослин, апікальні ділянки яких часто піддаються руйнуванню або пошкодженню, наприклад об’їдаються травоядними тваринами (у злаків) або ушкоджуються хвилями (у бурих водоростей); при цьому відпадає необхідність гілкування. |
Збільшення у довжину |
В ній розміщено кілька невеликих вакуолей і дрібні недиференційовані клітини – пропластиди.
Клітини апікальної меристеми мають назву ініціалі. При мітотичному поділі ініціалей одна з дочірніх клітин залишається у меристемі, а інша – збільшується за розміром, диференціюється і стає частиною постійного тіла рослини.
За походженням меристеми бувають первинні й вторинні. Первинна (апікальна) зумовлює розвиток проростка і первинний ріст органів (клітини зародкових стебла і кореня, що діляться). Вторинна меристема (латеральна) виникає з первинної.
Відомі два типи латеральної меристеми:
судинний камбій, що складається з клітин двох типів:
веретеноподібні ініціалі – клітини, здатні ділитися мітотично, утворюючи іззовні вторинну флоему, а всередині – вторинну ксилему;
променеві ініціалі – клітини майже сферичної форми, діляться мітотично, даючи початок паренхімним клітинам, які накопичуються і утворюють промені між сусідніми флоемою та ксилемою;
короковий камбій або фелоген – утворюють фелодерму або вторинний корок.
Культивування клітин рослин
Використання меристемних тканин є найпростішим способом вирощування нових рослин з використанням культур тканин
Калус – це недеференційована клітинна маса, здатна до росту і розмноження.
Клонуванням називають створення численних генетичних копій одного індивідуума за допомогою безстатевого розмноження. Процес цей може відбуватися природним шляхом, однак були також розроблені методи, що дозволяють проводити його штучно.
Клонування клітин з наступним їх скринінгом та регенерацією рослин з відібраних клонів розглядають як важливий метод збереження і поліпшення деревних порід помірних широт, зокрема хвойних дерев. Рослини-регенеранти, вирощені з клітин або тканин меристеми, використовують нині для розведення спаржі, суниці, брюссельської і кольорової капусти, гвоздик, папороті, персиків, ананасів, бананів тощо.
З клонуванням клітин пов'язують надії на усунення вірусних захворювань рослин. Розроблено методи, що дозволяють одержувати регенеранти з тканин верхівкових бруньок рослин (апікальної меристеми). Серед регенерованих рослин проводять відбір особин, вирощених з незаражених клітин, і вилучення хворих рослин, виявлених методом імунодіагностики.
Клонування клітин – перспективний метод одержання не тільки нових сортів, але і промислово важливих продуктів. За правильного підбору умов культивування, зокрема оптимального співвідношення фітогормонів, ізольовані клітини більш продуктивні, ніж цілі рослини. Іммобілізація рослинних клітин чи протопластів нерідко веде до підвищення їхньої синтетичної активності.
Комерційне значення в основному має промислове виробництво шиконіну. Застосування рослинних клітин, що є високоефективними продуцентами алкалоїдів, терпенів, різних пігментів і олій, харчових ароматичних добавок (суничної, виноградної, ванільної, томатної, селерової, спаржевої), наштовхується на певні труднощі, пов'язані з дорожнечею використовуваних технологій, низьким виходом цільових продуктів, тривалістю виробничого процесу.
28. Особливості структури та роль білків у життєдіяльності клітини.
Білки – найважливіша складова клітин будь-якого живого організму – не трапляються у неживій природі. Вони становлять значну частину тканин живого організму – до 25 % сирої та до 45 – 50 % сухої маси. Функції білків надзвичайно важливі та різноманітні. До них належать:
будівельна – білки утворюють основу цитоплазми та входять до складу всіх клітинних органоїдів;
каталітична – біологічні каталізатори називаються ферментами. Всі ферменти є білками, які каталізують біохімічні реакції. (Всі ферменти – білки, але не всі білки – ферменти);
рухова – будь-які форми руху у живій природі (робота м’язів, рух джгутиків у мікроорганізмів, рух цитоплазми) здійснюється білковими структурами клітини;
транспортна – білок гемоглобін транспортує кисень та СО2; білок альбумін – транспортує жирні кислоти тощо;
захисна – найважливіші чинники імунітету – антитіла – білки. До складу шкіри входить білок колаген; кератин – білок волосяного покриву.
регуляторна – ряд гормонів (речовин-регуляторів) є білками (інсулін);
запасна – білки здатні накопичуватися як запасні поживні речовини для харчування організму, що розвивається (овальбумін яйця, казеїн молока, білки насіння рослин тощо);
опорна – сухожилля, суглобні зчленування, кістки скелета, ратиці значною мірою складаються з білків.
Мономерами білків є амінокислоти. У білках міститься 20 різних амінокислот, які поділяються на незамінні, які можуть синтезуватися тваринними організмами з інших сполук і повинні надходити з їжею та замінні, що синтезуються у тваринному організмі у достатній кількості з незамінних амінокислот або інших сполук.
Окрім білків до складу всіх без винятку живих організмів входять нуклеїнові кислоти – генетичний матеріал будь-яких живих систем.
29. Ферменти, особливості їх структури, властивості та роль у процесах життєдіяльності клітини.
Ферменти (ензими) – клітинні каталізатори біохімічних реакцій, прискорюють їх, але самі у реакціях не витрачаються. Оскільки основа всіх ферментів – білкові молекули, які синтезуються у живій клітині, їх називають біологічні каталізатори. Вони каталізують численні реакції, які проходять при температурах, сумісних з життєвими процесами у клітині (5 – 40 С).
Речовина, перетворення якої каталізує фермент, має назву субстрат. З’єднуючись з субстратом, фермент утворює ферментсубстратний комплекс, імовірність перебігу реакції у якому значно зростає. Після закінчення реакції фермент вивільняється і знову може вступати у реакцію:
S + Е ES Û EP Û E + P, де
S – субстрат; Е – фермент; ЕS – ферментсубстратний комплекс; ЕР – комплекс фермент-продукт; Р – продукт (продукти).
Швидкість ферментативної реакції – кількість перетвореного субстрату або кількість продукту, утвореного за одиницю часу.
Властивості ферментів такі:
вони каталізують лише певні реакції (специфічність);
фермент просторово сполучається з речовинами, які вступають у реакції, прискорює їх перетворення і виходить з реакції незмінним;
присутність ферменту не впливає ні на природу, ні на властивості кінцевого продукту (продуктів) реакції;
ферменти – макромолекули – глобулярні білки. Речовини можуть бути низькомолекулярними. Тому активна не вся молекула ферменту, а лише її частина – активний центр, який геометрично відповідає структурі молекул речовин, які вступають в реакцію “ключ і замок”;
якщо відбувається денатурація білка, який є основою ферменту, то втрачається його активність, бо порушується структура його активного центра;
для ферментів у клітині й у організмі характерна певна локалізація, оскільки процеси розщеплення або синтезу теж мають певну локалізацію;
дія ферменту дуже ефективна (невелика кількість ферменту викликає перетворення великої кількості субстрату);
ферменти знижують енергію активації реакції, що каталізують.
Активність ферменту змінюється залежно від рН, температури, а також від концентрацій як ферменту, так і самого субстрату.
Вплив температури на активність ферментів. З підвищенням температури прискорюється рух молекул, внаслідок чого у молекул субстрату і ферменту є більше шансів на зіткнення і перебіг реакції. Температура, що забезпечує максимальну активність ферменту, має назву оптимальної температури ферментативної реакції (tоopt). Якщо, температура вище за tоopt, швидкість реакції знижується, оскільки відбувається руйнування вторинної і третинної структур білків, тобто інактивація ферменту.
Вплив рН на активність ферментів. При постійній температурі ферментативні реакції проходять найефективніше у вузьких межах рН. Оптимальне значення рН ферментативної реакції – те значення рН, при якому реакція відбувається з максимальною швидкістю. Зі зниженням рН збільшується концентрація іонів водню у середовищі, що веде до руйнування іонних зв’язків, що беруть участь у підтриманні просторової структури молекули ферменту. При різких змінах рН молекула ферменту може денатуруватися.
Вплив концентрації фермента на швидкість ферментативної реакції. За високої концентрації субстрату та сталих інших факторів (рН, tо) швидкість реакції пропорційна концентрації ферменту. Ферментативні реакцї відбуваються за умови, коли концентрація ферменту набагато нижча за концентрацію субстрату і тому з підвищенням концентрації ферменту швидкість реакції також зростає.
Вплив концентрації субстрату на швидкість ферментативної реакції. Для певної концентрації ферменту швидкість реакції збільшується зі збільшенням концнтрації субстрату, оскільки збільшується вірогідність контакту між ними. Але настає момент, коли збільшення концентрації субстрату вже не веде до збільшення швидкості реакції, оскільки всі активні центри ферменту зайняті.
30. Особливості структури і роль у клітині нуклеїнових кислот. Реплікація (самоподвоєння) ДНК
Нуклеїнові кислоти – найбільші з молекул живих організмів. Існують два види нуклеїнових кислот: ДНК – містить генетичну інформацію, куди входить інформація і про послідовність амінокислот у білках, та РНК, що бере участь у синтезі білків. Мономерами нуклеїнових кислот є нуклеотиди, що складаються з п’ятивуглецевого цукру (пентози), ортофосфорної кислоти та азотистої основи. ДНК містить дезоксирибозу, РНК – рибозу. Азотисті основи є похідними пурину та піримідину. До пуринових основ належать аденін (А) та гуанін (Г); до піримідинових – цитозин (Ц), урацил (У) та тимін (Т).
Реплікація ДНК. Існує кілька поглядів на те, яким чином відбувається процес реплікації ДНК. Запропоновано три найвірогідніші моделі проходження процесу. Вчені схиляються до напівконсерватиної.
Моделі
реплікації ДНК: 
а – напівконсервативна; б – консервативна; в – дисперсивна; 1 – материнські ділянки ДНК; 2 – синтезовані ділянки ДНК.
У процесі реплікації (самоподвоєння) ДНК певну роль відіграє принцип комплементарності.
Реплікація подвійної спіралі ДНК починається з того, що спіраль ДНК розкручується і відбувається розрив водневих зв’язків між комплементарними основами. Полінуклеотидні ланцюги не розриваються завдяки міцності їхніх фосфодиефірних зв’язків. Основи матричного ланцюга і вільні нуклеотидтрифосфати паруються, і останні утворюють полінуклеотидний ланцюг. Таким чином утворюються дві однакові молекули ДНК. У розділі ланцюгів і утворенні нового полінуклеотидного ланцюга бере участь фермент ДНК-полімераза (рис. 55). ДНК, подібно матриці, лише задає порядок розташування нуклеотидів у ланцюгах, що синтезуються, а сам процес веде фермент.
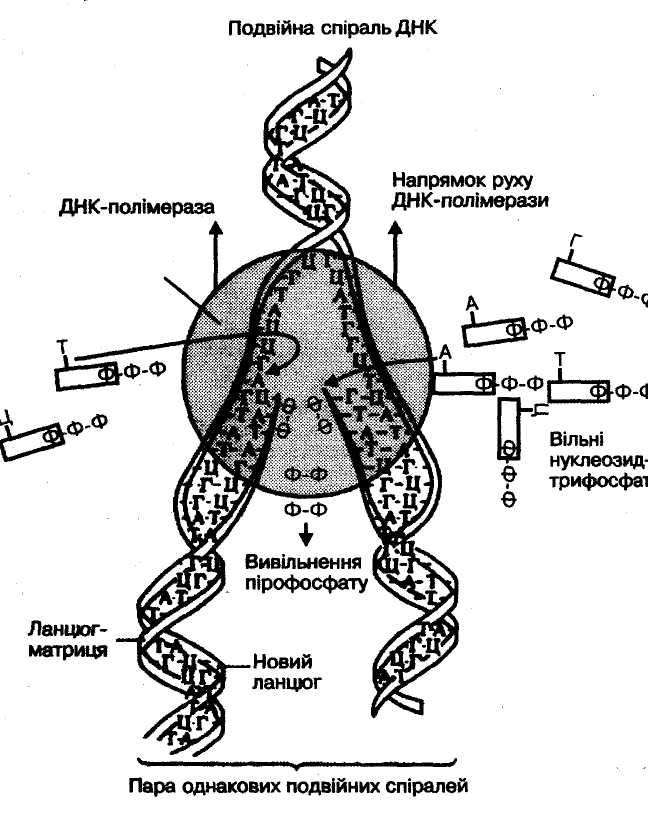
Схема реплікації ДНК
Різні досить довгі ділянки молекули ДНК відповідальні за синтез різних білків. Тим самим одна молекула ДНК може визначити синтез великого числа функціонально і хімічно різних білків клітини. За синтез кожного типу білків відповідальна певна ділянка молекули ДНК, яку часто позначають терміном "цистрон". На даний час поняття цистрон розглядають як еквівалентне поняттю ген. В унікальній структурі гена – у визначеному послідовному розташуванні його нуклеотидов уздовж ланцюга – закладена вся інформація про структуру одного відповідного білка.
Ген – одиниця спадкової інформації. Інформація одного гена відповідає структурі цілого поліпептидного ланцюга.
Зв’язок між основами у молекулі ДНК та амінокислотною послідовністю у молекулі білка має назву генетичного кода.
Властивості генетичного кода такі: він – триплетний: кожна з амінокислот, які входять до складу білка кодується послідовністю з трьох нуклеотидів – триплетом, який отримав назву кодон. Він також однозначний – кожний триплет кодує лише одну амінокислоту. Генетичний код не перекривається – зчитування генетичної інформації може відбуватися лише одним способом і неможливе іншим. Він вироджений – одна амінокислота може кодуватись не одним, а кількома певними триплетами. Генетичний код універсальний – він єдиний для всіх організмів, які існують на Землі. У прокаріотів, рослин, тварин, людини одні й ті самі триплети кодують одні й ті самі амінокислоти.
31. Етапи біосинтезу білків. Роль ДНК та РНК у біосинтезі білків. Відмінності біосинтезу білків у про- та еукаріотів.
Біосинтез білків - процес, за допомогою якого клітини будують білки. Термін іноді використовується для посилання виключно на процес трансляції, але частіше означає багатокроковий процес, що включає біосинтез амінокислот, транскрипцію, процесинг (включаючи сплайсинг), трансляцію та посттрансляційну модифікацію білків. Біосинтез білків, хоча й дуже подібний, дещо відрізняється між представниками трьох доменів життя —еукаріотами, археями та бактеріями.
Під час транскрипції відбувається зчитування генетичної інформації, зашифрованої в молекулах ДНК, і запис цієї інформації в молекули мРНК. Під час ряду послідовних стадій процесингу з мРНК видаляються деякі фрагменти, непотрібні в подальших стадіях (сплайсинг), і відбувається редагування нуклеотидних послідовностей. Після транспортування зрілої молекули мРНК з ядра до рибосом відбувається власне синтез білкових молекул шляхом приєднання окремих амінокислотних залишків до поліпептидного ланцюжка, що росте. На останній стадії посттрансляційної модифікації відбуваються зміни новосинтезованого білка додаванням небілкових молекул до білка та ковалентними модифікаціями його амінокислот.
Біосинтез білка проходить такі етапи:
активація – у цитоплазмі кожна амінокислота за участю ферментів приєднується міцним ковалентним зв’язком до відповідної молекули т-РНК;
ініціація – м-РНК, яка несе інформацію про синтез поліпептида, зв’язується з рібосомою, а потім з амінокислотою, прикріпленою до певної т-РНК; т-РНК, що несе амінокислоту, за принципом комплементарності взаємодіє з особливим триплетом (кодоном), який дає сигнал про початок синтезу поліпептидного ланцюга;
елонгація – подовження поліпептидного ланцюга за рахунок послідовного приєднання міцним ковалентним зв’язком амінокислот; кожна амінокислота займає своє положення за допомогою відповідної т-РНК, яка утворює комплементарні пари з визначеним кодоном у м-РНК;
термінація та звільнення – завершення синтезу поліпептидного ланцюга, про що сигналізує термінуючий кодон м-РНК (УАА, УАГ, УГА) і звільнення білка з рибосоми;
процесінг – згортання поліпептидного ланцюга, утворення у цитоплазмі просторової конфігурації молекули білка і утворення функціонально активної молекули білка (за допомогою ферментів відщеплюються ініціюючі амінокислоти та зайві амінокислотні залишки).


Молекула ДНК і окремі її функціональні ділянки, що несуть інформацію про структуру білків, самі безпосередньо участі в процесі створення білкових молекул не беруть. Першим етапом на шляху до реалізації цієї інформації, записаної в ланцюгах ДНК, є процес транскрипції, або "переписування".
Біосинтез білка складається з таких процесів:
транскрипція – передача інформації про структуру білка, закодованої в молекулі ДНК, на м-РНК (і-РНК): нитки ДНК за допомогою ферментів роз’єднуються і на одному з ланцюгів ДНК за принципом комплементарності синтезується молекула м-РНК (рис. 56);
трансляція – процес реалізації інформації про структуру білка, записаної в м-РНК у вигляді послідовності нуклеотидів, у послідовність амінокислот в молекулі білка.
32. Особливості структури та роль вуглеводів у життєдіяльності клітини.
Вуглеводи разом із білками – найпоширеніші сполуки, що беруть участь у побудові клітини та використовуються нею в процесі життєдіяльності. Поняття вуглеводи охоплює альдегіди та кетони багатоатомних спиртів, а також полімери цих сполук із загальною формулою Сn(H2O)m. У біосфері вуглеводів більше, ніж усіх органічних сполук у сукупності. У рослинному світі на них припадає 80 – 90 % всієї маси рослин. У тваринному організмі вміст вуглеводів становить близько 2 % маси тіла, проте їх значення для організму дуже велике.
Вуглеводи – продукти, які під дією енергії сонячного світла утворюються у фотосинтезуючих організмах з неорганічного вуглецю та води і дають початок іншим органічним сполукам живих організмів.
Функції вуглеводів важливі та різноманітні.
енергетична – окиснюючись у процесі клітинного дихання, вуглеводи вивільняють закладену в них енергію, забезпечуючи значну частину енергетичної потреби організму;
пластична – вуглеводи беруть участь у синтезі багатьох найважливіших для організму речовин: нуклеїнових кислот, амінокислот, ліпідів;
захисна – вуглеводи є основним компонентом оболонок рослинних клітин, беруть участь у побудові зовнішнього скелета членистоногих (хітиновий покрив);
опорна – целюлоза та інші полісахариди оболонок рослинних клітин і грибів не тільки захищають клітини від зовнішніх впливів, а й утворюють міцний стовбур (стебло, талом) організму, його механічні та опорні тканини.
Вуглеводи поділяють на моносахариди, олігосахариди та полісахариди.
Моносахариди – прості цукри (глюкоза, фруктоза, рибоза, дезоксирибоза). У молекулі – від трьох до дев’яти атомів вуглецю. Найпоширеніші – п’яти- і шестиатомні. Важливі як джерело енергії, а також як будівельні блоки для синтезу полісахаридів.
Олігосахариди – вуглеводи, які мають від 2 до 10 моносахаридів. Зв’язок між моносахаридами – глікозидний. Якщо моносахаридів – два, то це – дисахарид. Серед найвідоміших дисахаридів – лактоза (молочний цукор), цукроза (поширена у рослинах), мальтоза (продукт часткового гідролізу крохмалю).
Полісахариди – основна маса вуглеводів, представлених у природі, містять 10 і більше ланок моносахаридів. Ці сполуки, в основному – резерв поживних речовин та енергії (крохмаль та ґликоґен) і використовуються як будівельний матеріал (целюлоза). Крохмаль та ґлікоґен – полімери глюкози, останній – більш розгалужений.
33. Особливості будови та роль РНК у клітині. Типи РНК
РНК – одноланцюжковий (у більшості випадків) полінуклеотид.
Види РНК:
м-РНК (і-РНК) – матрична (інформаційна) РНК переносить інформацію про амінокислотну послідовність у будові білка з ДНК до місця синтезу – рибосом;
т-РНК – транспортна РНК переносить амінокислоти до рибосом;
р-РНК – рибосомальна РНК визначає структуру рибосом (разом з білками – їхній основний компонент).
34. Елементарний хімічний склад клітин мікроорганізмів. Роль макро-, оліго-, та мікроелементів у життєдіяльності клітини.
Елементний склад клітини можна поділити на чотири групи:
-
макроелементи (вміст яких від 1 % сухих речовин клітини та вище): органогенні H(6 – 8 %), O(20 – 25 %), N(7 – 14 %), C(46 – 50 %), P (3 %), Ca (1 %);
-
олігоелементи (від 0,1 – 1 %) K(1 %), Na, Cl, S (1 %), Mg(0,5 %), Fe(0,2 %) – зола;
-
мікроелементи (менше 0,1 %): Zn, Mn, Co, Cu, F, Br, I;
-
ультрамікроелементи – решта елементів; їхня концентрація в організмі дуже мала: 10-4 – 10-6 %; 12 з них визнано необхідними для життя рослин і тварин (B, Li, Al, Ni, Si, Sn, Cd, Ar, Se, Va, Ti, Cr).
Деякі незамінні мінеральні речовини та їх функції у клітині
|
|
|
|
Макроелементи |
|
|
С |
Синтез основних компонентів клітини |
|
О |
Синтез основних компонентів клітини |
|
N |
Синтез білків, нуклеїнових кислот, інших органічних сполук (коферменти, хлорофіл) |
|
Н |
Синтез основних компонентів клітини |
|
Р |
Синтез нуклеїнових кислот, АТФ, деяких білків, входить до складу біомембран |
|
S |
Синтез деяких білків (кератин), органічних сполук (кофермент А) |
|
К |
Бере участь у функціонуванні мембран: підтриманням електричного потенціалу, осмотичного та аніонно-катіонного балансу, проведенням нервових імпульсів. Кофактор ферментів, що беруть участь у фотосинтезі та диханні, компонент клітинного соку рослинних вакуолей |
|
Na |
Дія схожа на дію калію, але потреба у натрію значна менша |
|
Mg |
Входить до складу хлорофілу, клітинних стінок, мембран, кофактор для багатьох ферментів (АТФаза, кіназа) |
|
Са |
Зумовлює нормальний розвиток клітинної стінки, входить до складу екзоферментів (амілаз, протеїназ), у вигляді солі дипіколінової кислоти зумовлює термостійкість ендоспор бактерій |
|
Cl |
Підтримання осмотичного аніонно-катіонного балансу |
|
Fe |
Перенесення електронів фосфатних груп, окиснення жирних кислот, дихання, фотосинтез |
|
Со |
Міститься у коферменті вітамін-В12-ферментів |
|
Сu |
Перенесення електронів у дихальному ланцюгу, каталізує перетворення кисню на воду |
|
Zn |
Міститься у ферментах алкогольдегідрогеназі (анаеробне дихання), карбоксипептидазі тощо. |
|
Мо |
Міститься у ферментах нітрогеназі (фіксація азоту), нітратредуктазі (відновлення NO3- до NO2-) |
35. Метаболізм клітини. Значення АТФ у процесах анаболізму та катаболізму.
Обмін
речовин (метаболізм) живої клітини
складається з двох протилежно спрямованих
видів реакцій – катаболічних і
анаболічних.
Фн – ортофосфорна кислота
Катаболізм (енергетичний обмін) – сукупність реакцій розпаду органічних сполук. Біополімери (білки, полісахариди та ліпіди), залучаються до катаболічних реакцій у три стадії.
На першій стадії катаболізму великі молекули полімерів розпадаються на мономерні субодиниці. Цей процес, що називається травленням і відбувається поза клітинами під дією травних ферментів.
На другій стадії невеликі утворені молекули надходять до клітини організму і зазнають подальшого розщеплення у цитоплазмі. При цьому велика частина всіх мономерів перетворюється на піровиноградну кислоту (піруват), а потім – на ацетилкофермент А (рис. 52).
Остання, третя стадія катаболізму полягає у повному розщепленні ацетильної групи ацетил коферменту А до СО, та Н2О, що супроводжується утворенням АТФ з АДФ та фосфорної кислоти:
АДФ + Н3РО4 = АТФ + Н2О.
Третя стадія катаболізму можлива тільки за наявності вільного кисню, тобто проходить винятково в аеробних організмах, і для її здійснення необхідні біомембрани. В еукаріотичних клітинах ця стадія проходить у мітохондріях. У анаеробів, у безкисневих умовах, катаболізм.
Такі процеси називаються бродінням (молочнокисле, спиртове), а процес перетворення простих цукрів на піруват — ґліколізом.
Перевага аеробів перед анаеробами пов’язана з тим, що основна кількість енергії вивільняється з поживних речовин саме на третій стадії катаболізму, якої нема у анаеробів.
Розщеплення ацетильної групи ацетилкоферменту А відбувається у мітохондріях у циклі лимонної кислоти (цикл Кребса). Реакції циклу поставляють редукційний потенціал у систему транспорту електронів, сполучену з синтезом АТФ. Таким чином мітохондрії виконують свою головну функцію — генераторів енергії у клітині.
Всі описані процеси (ґліколіз, цикл лимонної кислоти, синтез АТФ та ін.) становлять основу енергетичного обміну, тобто біохімічних реакцій, що забезпечують накопичення енергії у вигляді молекул АТФ. Реакції синтезу органічних сполук, або пластичний обмін, проходять із витратою енергії. При цьому створюються необхідні для організму біополімери.
Катаболізм та анаболізм постійно пов’язані між собою потоками речовин та енергії. У катаболічних і анаболічних процесах беруть участь одні й ті самі молекули, які утворюють потік речовин. Потік енергії представлений зворотним процесом синтезу — розпаду АТФ.
Анаболізм (пластичний обмін) – сукупність реакцій синтезу органічних сполук. Найважливішими з анаболічних реакцій є реакції матричного синтезу.
Обмін речовин виконує функції забезпечення клітини: будівельним матеріалом в результаті пластичного обміну, що забезпечує побудову клітини та оновлення її складу; енергією в результаті енергетичного обміну. Обмін речовин – зв’язок із зовнішнім середовищем. Це – основні умови підтримання життя клітини. Пластичний та енергетичний обміни тісно пов’язані.