

7.2.4. Взаимодействие ауксинов и цитокининов
7.2.4.1. Физиологическое действие ауксинов и цитокининов в культуре in vitro
С открытием цитокининов началось культивирование растительных клеток in vitro. Первым типом ткани, полученным из паренхимы табака, был каллус. В природе каллусы образуются в местах повреждения: растению необходимо быстро зарастить шрам, заполняя его бесформенной массой клеток. Поврежденная проводящая система (сосуды), покровные и механические ткани восстанавливаются позже. Клетки каллуса быстро делятся, веретено деления располагается в случайном направлении. При этом получается рыхлая клеточная масса.
Для стимуляции деления клеток in vitro в среду добавляют и ауксины, и цитокинины. Показано, что ауксины активируют CDK-протеинкиназы клеточного цикла (cycline dependent kinases), а цитокинины — соответствующие циклины. Комплекс CDK-циклин необходим для запуска клеточного деления. Изменение соотношения ауксин/цитокинин приводит к морфогенезу in vitro. При преобладании ауксинов начинается ризогенез (от греч. rhiza — корень; genesis — рождение). Если же преобладают цитокинины, то образуются меристемы побегов, т.е. начинается геммагенез (от греч. gemma — почка) (рис. 7.13).
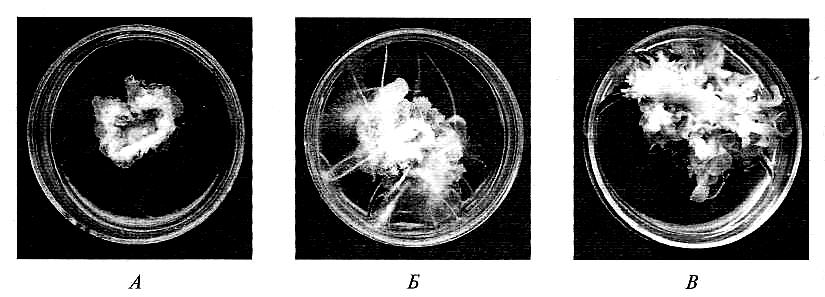
Рис. 7.13. Действие ауксинов и цитокининов на каллусную ткань:
А — физиологическая активность ауксинов и цитокининов приблизительно равны — развивается каллусная ткань; Б — ауксины преобладают над питокининами — ризогенез (образование корней); В — цитокинины преобладают над ауксинами — геммагенез (образование побегов)
Такое поведение культуры клеток хорошо согласуется с функцией ауксинов и цитокининов как «гормонов благополучия» побегов и корней соответственно. Недостаток ауксинов воспринимается клетками как недостаточное развитие побегов и служит сигналом для их образования. В дифференцированных побегах синтезируется ауксин и баланс гормонов восстанавливается. Аналогичный механизм работает при недостатке цитокининов, тогда формируются корни. При удалении из среды и ауксинов, и цитокининов в культуре клеток иногда образуются биполярные структуры — эмбриоиды (embryo — зародыш; eidos — похожий). У каждого эмбриоида есть свой источник цитокининов (корневой полюс) и ауксинов (побеговый полюс). Культуру клеток используют в технологиях ускоренного размножения растений.
7.2.4.2. Баланс между ауксинами и цитокининами в интактном растении
Итак, для нормальной жизни в растении всегда должен существовать баланс между ауксинами и цитокининами. Процесс вегетативного роста растений можно моделировать, рассматривая только полярный транспорт и взаимодействие ауксинов и цитокининов. Если главный стебель отделить от корня, то в верхней части останется точка синтеза ауксинов и возникнет дефицит цитокининов. ИУК из апекса транспортируется к основанию черенка, где создается избыток ауксинов. Для восстановления баланса нужны цитокинины, т. е. формируются корни. У оставшейся нижней части растения есть точки синтеза цитокининов, но не хватает ауксинов. Цитокинины транспортируются вверх и накапливаются около среза. Это также вызывает нарушение баланса и ведет к синтезу ауксинов: пробуждаются спящие почки или из каллусной массы возникают новые побеги.
В интактном растении активные деления клеток происходят в апексах корня и побега. Согласно модели, чем дальше находится орган от кончика корня, тем меньше в нем цитокинина. Апексом побега транспорт заканчивается, и происходит накопление цитокинина. Ауксин синтезируется в апексе побега. Таким образом, в меристеме побега концентрация цитокининов и ауксинов оказывается достаточной для поддержания клеточных делений.
Обратная ситуация в апексе корня. Ауксин накапливается, так как апекс корня — конечный пункт транспорта ИУК. Вместе с синтезируемыми в корне цитокининами ауксины вызывают клеточные деления в меристеме корня. Чем больше расстояние от кончика корня до верхушки побега преодолевают гормоны, тем меньше их содержание в меристеме. Наконец, наступает момент, когда цитокининов, поступающих в апекс побега, недостаточно для делений. Рост стебля вверх останавливается. Аналогично регулируется рост корня вглубь. Таким образом, полярный рост растения можно моделировать на основании баланса ауксинов/цитокининов.
Расположение камбия в растении также не случайно. По флоэме вниз движется ауксин, а по ксилеме вверх — цитокинин. Между ними на «перекрестке» потоков ауксинов и цитокининов (а также фотоассимилятов и минеральных веществ), необходимых для деления, находятся делящиеся клетки камбия. Работа камбия также моделируется исходя из теории баланса фитогормонов: если ауксин преобладает, камбий откладывает больше ксилемы, обеспечивая приток цитокининов из корней, избыток цитокининов ведет к закладке флоэмы (по ней поступят новые порции ауксина, снова возникнет равновесие между двумя типами гормонов). Поврежденные корни и ветви утолщаются гораздо хуже. Причина этого — в низкой способности к синтезу гормонов. Однако чем больше потоки гормонов через ветку или корень, тем сильнее работа камбия, больше флоэмы и ксилемы, выше механическая прочность.