

7.2.1.3. Фосфатидилинозитольная система вторичных мессенджеров
Принципы работы вторичных мессенджеров в данном подразделе показаны на примере фосфатидилинозитольной системы (рис. 7.3). Фосфатидилинозитол — один из липидов плазмалеммы, поэтому фосфатидилинозитольная система работает с рецепторами, расположенными на внешней мембране.
При связывании с гормоном белок-рецептор меняет конформацию и передает сигнал на G-белок. G-белки названы так за способность связывать и гидролизовать ГТФ. Конформация белкового комплекса рецептора влияет на способность G-белка связывать и удерживать ГТФ.
Пока молекула ГТФ остается связанной с G-белком, он успевает активизировать до 10 молекул фосфолипазы С, входящей в состав мембранных белков. После этого происходит гидролиз ГТФ, ГДФ и фосфат освобождаются, G-белок переходит в неактивное состояние, пока не захватит новую молекулу ГТФ. Работа G-белка регистрируется в первые 1 — 5 с после обработки рецептора гормоном (рис. 7.3, Б).
Активизация фосфолипазы С приводит к гидролизу фосфатидилинозитолдифосфата. При этом в мембране остается гидрофобный остаток диацилглицерина, а в цитоплазму поступает инозитол-1,4,5-трифосфат (ИФ3). Именно это событие и дало название всему каскаду вторичных мессенджеров. Уменьшение доли фосфатидилинозитолтрифосфата и поступление инозитолтрифосфата в цитоплазму наблюдается уже через 10 —15 с после получения сигнала, достигая максимума к 60 с.
Инозитолтрифосфат может взаимодействовать с Са2+-каналами на мембране. Через 2 мин после обработки гормоном в цитоплазму начинает поступать кальций. Этот процесс имеет характер осцилляции (так называемые «кальциевые волны»). Концентрация кальция в клетке изменяется приблизительно с 1 до 100 ммоль/л. Дальнейшее повышение концентрации кальция может вызвать автолиз клетки: активизируются Са2+-зависимые фосфолипазы (например, фосфолипаза А), что приведет к разрушению мембраны. Чтобы избежать автолиза, клетка модулирует уровень кальция через концентрацию инозитолтрифосфата. Так, при превышении уровня ИФ3 может происходить дополнительное фосфорилирование, а инозитолтетрафосфат (ИФ4) уже не взаимодействует с
Са2+-каналами. При этом инозитолтетрафосфат используется как депо, из которого можно «изъять» новую порцию активного инозитолтрифосфата. После достижения максимума инозитолтрифосфат поэтапно гидролизуется до инозитола. Далее инозитол включается в состав фосфатидилинозитола, и только в составе мембраны фосфорилируется до фосфатидилинозитолди-фосфата и используется еще раз. Фосфатидилинозитолдифосфат является важным модулятором при взаимодействии мембран с цитоскелетом. Так, повышение его доли в составе мембран провоцирует сборку актиновых филаментов, а дефосфорилирование приводит к разборке цитоскелета.
Инозитол в растительных клетках может полностью фосфорилироваться, превращаясь в инозитолгексафосфат (фитин). Обычно это соединение рассматривают как депо фосфора в семенах и запасных тканях. У дрожжей инозитолгексафосфат необходим для транспорта РНК из ядра в цитоплазму. Возможно, похожие явления будут обнаружены и у растений.
Кроме фосфатидилинозитольной системы у растений обнаружены еще несколько сигнальных систем. Например, аденилатциклазная система похожа по действию на аналогичную систему животной клетки. В липоксигеназной системе в регуляцию сигнального каскада вовлечена также фосфолипаза А, активируемая кальцием. После ее активизации в мембране появляются свободные жирные кислоты. Кроме того, кальций активизирует липоксигеназу, которая включает кислород в состав полиненасыщенных жирных кислот с образованием пятичленных углеродных колец. Одним из результатов работы липоксигеназы является синтез жасминовой кислоты, блокирующей Са2+-каналы, что предотвращает избыточное поступление кальция.
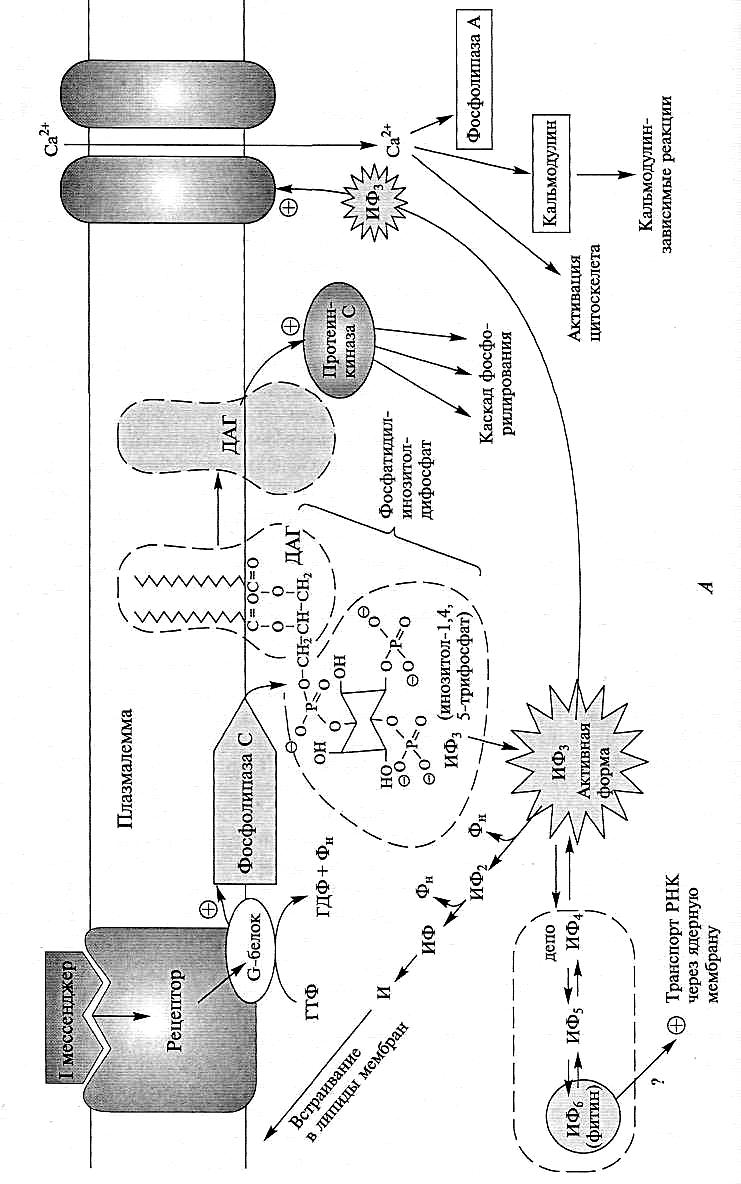
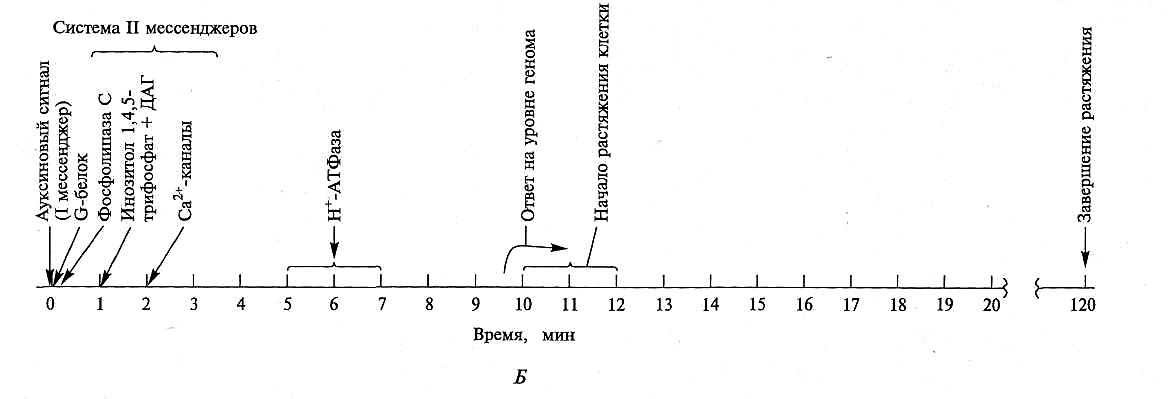
Рис. 7.3. Фосфатидилинозитольная система вторичных мессенджеров:
А — последовательность активизации вторичных мессенджеров в пространстве; Б — последовательность событий на шкале времени. И — инозитол; ИФ — инозитолфосфат; ИФ2 — инозитолдифосфат; ИФ3 — инозитолтрифосфат; ИФ4 — инозитолтетрафосфат; ИФ5 — инозитолпентафосфат; ИФ6 — инозитолгексафосфат (фитин); ДАГ — диацилглицерин
Есть сведения о работе в растительной клетке NO-системы, системы фосфатидной кислоты, НАДН-оксидазной системы, пероксидазной системы, МАР-киназного каскада (Mitogene activated proteinkinase).
Следующими по времени активируются функциональные системы клетки, происходит активация/инактивация генов, изменяется мембранный потенциал. Так, активация фосфатидилинозитольной системы ауксином вызывает выброс протонов из цитоплазмы за счет активизации Н+-помпы. Через 5 — 7 мин после воздействия гормона наблюдается закисление матрикса клеточной стенки. Ответ этого компартмента на закисление зависит от состава клеточной стенки, в частности от присутствия кислых гидролаз, инвертазы, доли пектиновых веществ и т.д. Увеличение размеров клетки начинается через 10 —12 мин после обработки ауксином.
При закислении клеточной стенки из пектина освобождается кальций, связанный с кислотными группировками. Таким образом, растительная клетка использует клеточную стенку как депо Са2+. Уровень Са2+ в цитоплазме зависит от концентрации инозитолтрифосфата и жасминовой кислоты, состояния матрикса клеточной стенки, работы Са2+-каналов и Са2+-АТФаз. Эти факторы действуют не одновременно, причем приток кальция в клетку провоцирует процессы, приводящие к снижению его концентрации и наоборот. В результате поток кальция оказывается неравномерным во времени, возникают волны поступления/удаления кальция. Динамика волн кальция существенно различается для разных внешних сигналов (см. подразд. 6.3.4.3). До 70 % поступившего в клетку кальция связывается с цитоскелетом, активизируя его работу.
Важным вторичным мессенджером в передаче сигнала являются небольшие белки — калъмодулины. Кальмодулин имеет 4 центра, связывающихся с Са2+, и его конформация различается в зависимости от числа занятых кальцием центров. От активности кальмодулинов, в частности, зависит работа Са2+-АТФаз: полностью заполненный кальцием кальмодулин усиливает выкачивание кальция из клетки. Гидрофобный диацилглицерин также является вторичным мессенджером. Он взаимодействует с протеинкиназой С, повышая ее фосфорилирующую активность. Далее запускается каскад фосфорилирования белков. Вторичными мессенджерами в приведенном примере выступают G-белки, фосфолипазы С и А, инозитолтрифосфат, жасминовая кислота, кальций, диацилглицерин, протеинкиназа С. При добавлении в клетку какого-либо из этих мессенджеров даже без первичного сигнала можно наблюдать физиологический ответ (обычно такой ответ бывает неполным).