

7.2.5.6. Гиббереллин и цветение растений
Гиббереллины стимулируют цветение ряда растений. Обычно уровень эндогенных ГК повышается при увеличении длины дня. У многих растений цветение контролируется фотопериодом. Виды, цветущие на длинном дне, можно заставить цвести с помощью гиббереллинов, В опытах М.Х.Чайлахяна к ГК оказались чувствительными рудбекия (Rudbeckia), каланхоэ, морковь (Daucus carota), другие виды не цвели. Участие гиббереллина в регуляции цветения очевидно, хотя результат зависит от частной физиологии объекта (см. подразд. 7.4.2).
7.2.6. Абсцизовая кислота — сигнал водного стресса
7.2.6.1. Открытие абсцизовой кислоты
В 1949 г. были проведены эксперименты с покоящимися почками березы. Водный экстракт почек замедлял рост гипокотилей, а при нанесении совместно с ауксинами ингибировал растяжение. Когда покой почек заканчивался, экстракт не угнетал рост. Исследователи предположили, что в почках содержится ростовой регулятор — дормин (от англ. dormancy — покой).
В 1962—1964 гг. в США Дж. Лью и Г. Р. Карнс изучали созревание и опадение коробочек хлопчатника (Gossypium hirsutum). Они предположили, что в хлопчатнике накапливается вещество, стимулирующее обезвоживание плода, его растрескивание и формирование отделительного слоя. Гипотетический гормон назвали абсцизином (от лат. abscisio — опадение). В 1965 г. абсцизин был выделен и были изучены его основные свойства. Выяснилось, что абсцизин и дормин — одно и то же соединение. Из-за приоритета сегодня установилось название, данное Лью и Карнсом, — абсцизовая кислота (АБК). Заметим, что АБК редко регулирует опадение плодов и листьев (обычно за это отвечает этилен: см. подразд. 1.2.1 Л). АБК нужна для регуляции покоя, поэтому название «дормин» гораздо лучше отражает суть регулируемых процессов.
7.2.6.2. Биосинтез абсцизовой кислоты
АБК является изопреноидом с 15 атомами углерода, соединенными в изопреновые С5-звенья (см. подразд. 9.6.2). Обычно изопреноиды синтезируются в пластидах из общего предшественника — изопентенилпирофосфата. В ранних работах было высказано предположение, что АБК синтезируется сразу после конденсации трех С5-звеньев, т.е. эта гипотеза предполагает прямой синтез АБК из более простых предшественников. В дальнейшем прямой синтез АБК был показан только для фитопатогенных грибов (для возбудителя серой гнили Botrytis).
Гипотеза прямого синтеза доминировала до 1990-х гг., пока не выяснилось, что у мутантов по биосинтезу каротиноидов нарушен и биосинтез АБК. На сегодня получены мутации по ферментам почти всех этапов биосинтеза АБК. Таким образом, АБК синтезируется через более сложные предшественники, можно рассматривать ее как продукт специфической деградации каротиноидов.
Первые этапы биосинтеза АБК связаны с ксантофилловым циклом (см. подразд. 3.3.4.4). Зеаксантин через промежуточный продукт антераксантин превращается в все-транс-виолаксантин. Дальнейшие превращения включают 9-цис-изомеризацию каротиноидного предшественника (на эту роль претендуют виолаксантин и неоксантин). В конечном итоге 9-цис-предшественник расщепляется на два неравных фрагмента: С15 (ксантоксин) и С25. С25-фрагмент быстро деградирует, а ксантоксин превращается в абсцизовый альдегид (рис. 7.16). Для последнего этапа биосинтеза необходим молибденсодержащий белок, катализирующий окисление АБ-альдегида до АБК. Молибденовый кофактор этого фермента общий с нитратредуктазой и ксантиндегидрогеназой. При мутациях по синтезу молибденового кофактора растение не может восстанавливать нитрат и окислять АБ-альдегид.
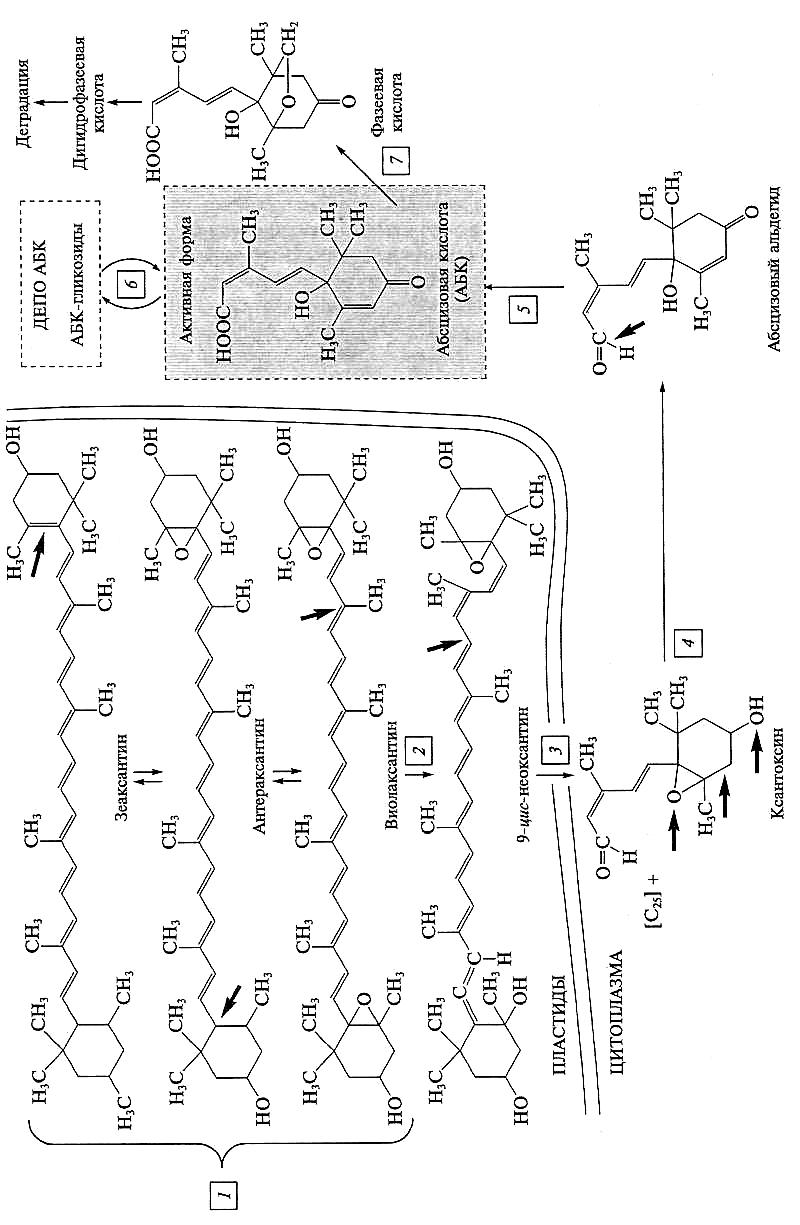
Рис. 7.16. Биосинтез, инактивация и деградация АБК:
1 — виолаксантиновый цикл в пластидах; 2 — ключевой этап биосинтеза АБК — цыс-изомери-зация каротиноидов; 3 — расщепление каротиноидов на С15- и С25-фрагменты и выход ксантокси-на в цитозоль; 4 — превращение ксантоксина в абсцизовый альдегид, стрелками показаны связи в молекулах, которые претерпевают модификацию на каждом этапе; 5 —окисление АБ-альдегида до физиологически активной АБК; 6 — депонирование АБК; 7 — необратимая деградация АБК