5. Анаболизм
Анаболизм – это биосинтез белков, полисахаридов, липидов, нуклеиновых кислот и других макромолекул из малых молекул-предшественников. Поскольку он сопровождается усложнением структуры, то требует затрат энергии. Источником такой энергии является энергия АТФ.
Также для биосинтеза некоторых веществ (жирные кислоты, холестерол) требуются богатые энергией атомы водорода – их источником является НАДФН. Молекулы НАДФН образуются в реакциях окисления глюкозо-6-фосфата в пентозном пути и оксалоацетата малик-ферментом. В реакциях анаболизма НАДФН передает свои атомы водорода на синтетические реакции и окисляется до НАДФ. Так формируется НАДФ-НАДФН-цикл.
Катаболизм
Катаболизм – расщепление и окисление сложных органических молекул до более простых конечных продуктов. Оно сопровождается высвобождением энергии, заключенной в сложной структуре веществ. Большая часть высвобожденной энергии рассеивается в виде тепла. Меньшая часть этой энергии "перехватывается" коферментами окислительных реакций НАД и ФАД, некоторая часть сразу используется для синтеза АТФ.
Следует заметить, что атомы водорода, высвобождаемые в реакциях окисления веществ, могут использоваться клеткой только по двум направлениям:
-
на анаболические реакции в составе НАДФН.
-
на образование АТФ в митохондриях при окислении НАДН и ФАДН2.
Амфиболические пути выполняют сразу несколько функций, они находятся на <перекрестках> метаболизма и связывают анаболические и катаболические пути; примером может служить цикл лимонной кислоты .
Цитратный цикл: реакции
В цитратном цикле (цикл лимонной кислоты; метаболический процесс, протекающий в матриксе митохондрий) ацетильные остатки (CH3CO—) окисляются до диоксида углерода (CO2). Полученные при этом восстановительные эквиваленты переносятся на НАД+ или убихинон и включаются в дыхательную цепь (см. с. 142). Центральная роль цитратного цикла в метаболизме клетки рассматривается на с. 140.
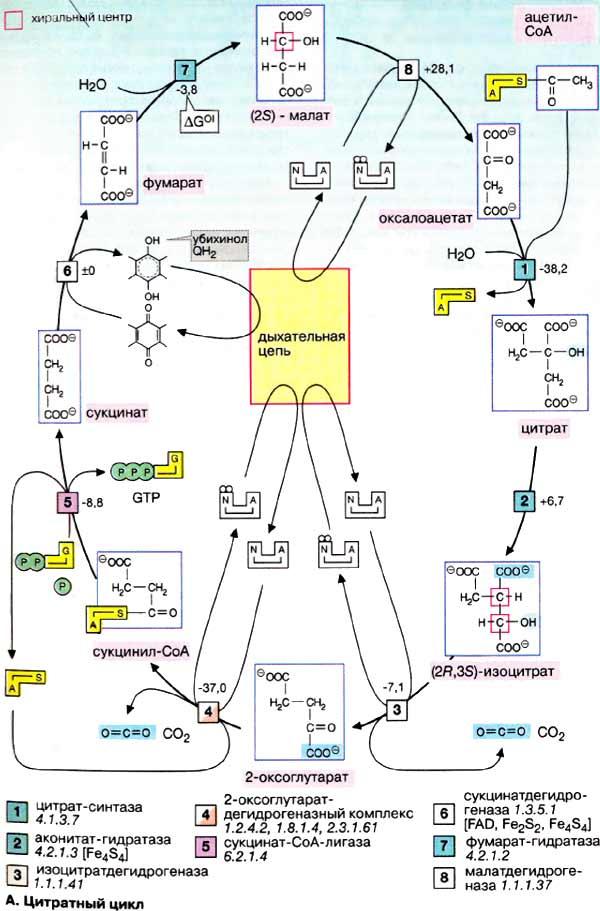
А. Цитратный цикл
Большая часть потребляемого в цитратном цикле ацетил-КоА получает ацетильные остатки, образовавшиеся в результате β-окисления жирных кислот (см. с. 166) и окислительного декарбоксилирования пирувата, катализируемого пируватдегидрогеназой (см. с. 136). Оба процесса протекают в матриксе митохондрий.
Окисление ацетильных остатков включает ряд промежуточных стадий, образующих цикл: сначала ацетильная группа в реакции, катализируемой цитрат-синтазой [1], конденсируется с молекулой оксалоацетата с образованием цитрата(цикл получил свое название по продукту этой реакции). На следующей стадии [2] цитрат изомеризуется в изоцитрат с переносом гидроксильной группы внутри молекулы. При этом промежуточный продукт реакции, ненасыщенный аконитат, остается во время реакции связанным с ферментом (на схеме не показано). Поэтому фермент, катализирующий реакцию, называют аконитат-гидратазой [2] («аконитазой»).
Свойства аконитат-гидратазы обеспечивают абсолютную стереоспецифичность изомеризации. В то время как цитратне обладает хиральностью, изоцитрат содержит два асимметрических центра и может существовать в четырех изомерных формах. Однако в цитратном цикле образуется только один из стерео изомеров, (2R,3S)-изоцитрат (см. с. 16).
На следующей стадии изоцитратдегидрогеназа (3) окисляет гидроксигруппу изоцитрата в оксогруппу с одновременным отщеплением одной из карбоксильных групп в виде CO2 и образованием 2-оксоглутарата. Последующее образование сукцинил-КоА [4], включающее реакции окисления и декарбоксилирования, катализируется мультиферментным комплексом, 2-оксоглутаратдегидрогеназой (дегидрогеназы кетокислотрассмотрены на предыдущей странице). Расщепление тиолсложноэфирной связи сукцинил-КоА с образованием сукцината и кофермента А, катализируемое сукцинат-КоА-лигазой («тиокиназой» (5)], — высокоэкзоэргическаяреакция, энергия которой используется для синтеза фосфоангидридной связи («субстратного фосфорилирования», см. с. 126). В цитратном цикле синтезируется не АТФ, как в большинстве таких реакций, а гуанозинтрифосфат [ГТФ (GTP)], который легко превращается в АТФ нуклеозиддифосфаткиназой (на схеме не показано).
В приведенных реакциях ацетильный остаток полностью окисляется до CO2. Однако одновременно молекулапереносчика оксалоацетата восстанавливается в сукцинат. В трех последующих реакциях цикла сукцинат снова превращается в оксалоацетат. Вначале сукцинатдегидрогеназа [6] окисляет сукцинат в фумарат. В отличие от других ферментов цикла сукцинатдегидрогеназа является интегральным белком внутренней митохондриальной мембраны. Поэтому ее относят также к комплексу II дыхательной цепи. Сукцинатдегидрогеназа содержит ФАД (FAD) в качестве простетической группы, однако фактическим акцептором электронов является убихинон. Затем к двойной связифумарата с помощью фумарат-гидратазы («фумаразы», [7]) присоединяется вода и образуется хиральный (2S)-малат. На последней стадии цикла малат окисляется малатдегидрогеназой (8) в оксалоацетат с образованием НАДН + Н+. Эта реакция замыкает цитратный цикл.
Общий баланс цитратного цикла состоит в том, что из одного ацетильного остатка образуются 2 CO2, 3 НАДН + Н+ и одна молекула восстановленного убихинона (QH2). При этом за счет восстановленных форм коферментов путемокислительного фосфорилирования в клетке синтезируются 9, а с учетом трансформации одной молекулы ГТФ — 10 молекул АТФ.
6.
ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ - это синтез АТФ из аденозиндифосфата и неорганического фосфата, осуществляющийся в живых клетках, благодаря энергии, выделяющейся при окислении органических веществ в в процессе клеточного дыхания.
СУБСТРАТНОЕ ФОСФОРИЛИРОВАНИЕ - это синтез АТФ, не связанный с электрон-транспортной системой, при котором остаток фосфорной кислоты (Н2РО3) переносится на АДФ от высокоэнергетического (фосфорилированного) соединения. Для ряда анаэробов (осуществляющих брожение) является единственным способом получения энергии.
В процессе биологического окисления около 50% энергии резервируется клетками тканей в макроэргических соединениях, преимущественно АТФ. Синтез АТФ из АДФ и фосфорной кислоты, который происходит с использованием энергии, выделяющееся при окисления веществ в живых клетках, и связанный с переносом электронов по дыхательной цепи, называется окислительным фосфорилированием.
Окислительного фосфорилирования может осуществляться на уровне субстрата (субстратное фосфорилирование), но главным образом на различных этапах дыхательной цепи. Субстратное фосфорилирование, как отмечалось выше, происходит путем непосредственной передачи молекулы активного фосфата с субстратов, содержащих макроэргических связь, на АДФ с образованием АТФ (см. Обмен углеводов, липидов). Например, промежуточный продукт распада глюкозы и триацилглицеры-нов 2-фосфоенолпировиноградна кислота отдает свой активный фосфат на АДФ с образованием АТФ за реакцией. Однако субстратное фосфорилирование дает незначительное количество молекул АТФ. Основное их количество синтезируется в процессе фосфорилирования, которое связано с клеточным дыханием. Установлено, что на каждом этапе переноса электронов от одного переносчика на другой они переходят с одного энергетического уровня на другой (ниже), в результате чего происходит высвобождение определенного количества энергии. Однако существует три этапа, когда энергии, высвобождаемой, достаточно для синтеза АТФ
На основе данных термодинамики допускала наличие трех участков (пунктов) дыхательной цепи, которые сопровождались синтезом АТФ. Опыты с применением специфических ингибиторов определенных ферментов дыхательной цепи подтвердили эти данные. Так, ро-тенон (инсектицид - токсичное вещество растительного происхождения, применяется индейцами как яд) блокирует перенос электронов на участке от НАДН2 к КОО. При этом все компоненты дыхательной цепи переходят в окисленный состояние, т.е. уменьшается скорость транспорта электронов. Амитал (барбитураты натрия) препятствует восстановлению КОО. Антибиотик антимицин А блокирует перенос электронов от цитохрома b цитохром Cj, а цианиды, азид натрия, сероводород связываются с цитохромоксы-ГАЗО и препятствуют переходу электронов с ЦХО на молекулярный кислород.
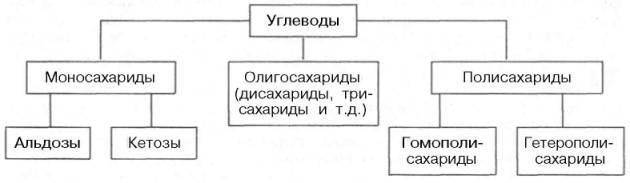
КЛАССИФИКАЦИЯ УГЛЕВОДОВ
Углеводы можно определить как альдегидные или кетонные производные полиатомных (содержащих более одной ОН-группы) спиртов или как соединения, при гидролизе которых образуются эти производные.
Согласно принятой в настоящее время классификации, углеводы подразделяются на три основные группы: моносахариды, олигосахариды и полисахариды.