

3.2. Механизмы действия липофильных гормонов
Липофильные гормоны, за исключением ЙТ и ретиноата, являются производными холестерина. После секреции свободный гормон диффундирует сквозь плазматическую мембрану и связывается с рецептором либо в гиалоплазме, либо в ядре. Возникший гормон-рецепторный комплекс связывается со специфическим участком ДНК, который называется гормон-реактивным элементом (ГРЭ). Он функционально аналогичен энхансеру или сайленсеру, и в результате меняется экспрессия гена (то есть активируются или ингибируются специфические гены). Например, происходит увеличение матки во время беременности, рост молочной железы перед и во время лактации.
Между двумя группами липофильных гормонов есть существенные различия в локализации их рецепторов (рис.11). Рецепторы стероидных гормонов находятся в цитозоле в неактивном комплексе с шаперонами, после их диссоциации гормон-рецепторный комплекс перемещается в ядро и там взаимодействует с ГРЭ. Рецепторы йодтиронинов и активных форм витаминов D (кальцитриол) и А (ретиноат) постоянно находятся в ядре клетки. Свободному гормону приходится проникать не только в клетку, но и в ядро и взаимодействовать с соответствующим ГРЭ ДНК.

Рис.11. Механизм действия гормонов липофильной природы
Вторая группа гормонов (гидрофильные) взаимодействует с рецептором на наружной стороне плазматической мембраны. В этом механизме различают:
быстрые системы, сопряженные с ионными каналами;
медленные системы, включающие вторые посредники и (или) протеинкиназы.
3.3. Быстрые системы
Быстрые системы опосредуют эффекты медиаторов, передающих нервный импульс за миллисек. Такую скорость может обеспечить только открытие ионных каналов (рис.12), приводящее к быстрому движению ионов от высоких концентраций к низким(вход Na+ и Сl- в клетку).
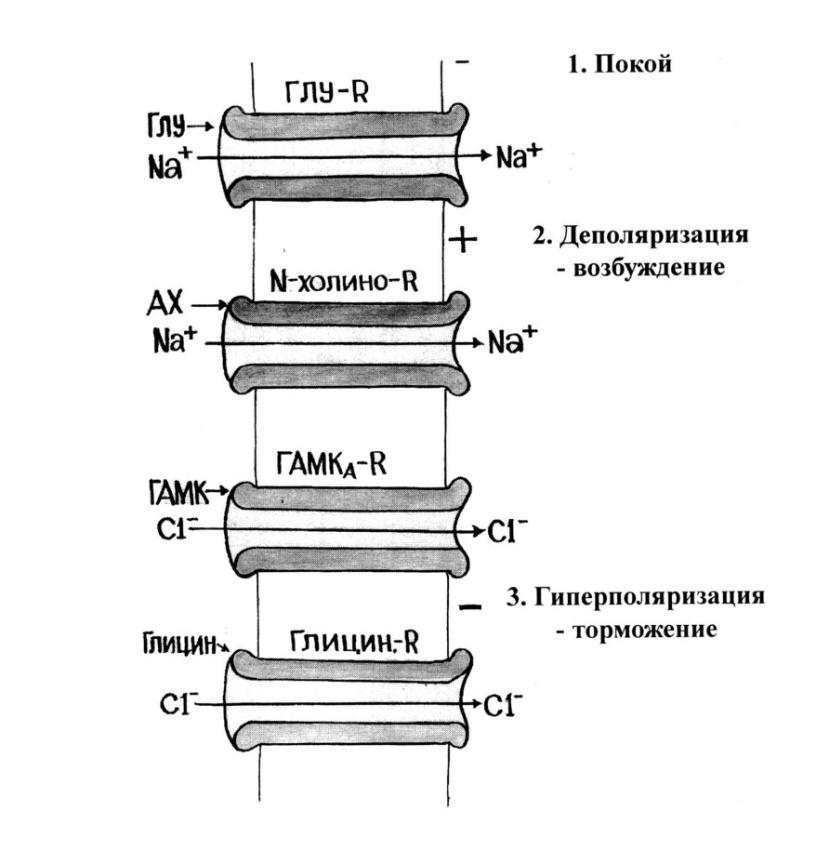
Рис.12. Механизм действия медиаторов
В состоянии покоя закрыты любые каналы, на плазматической мембране снаружи плюс, а на внутренней стороне – минус. При действии возбуждающей аминокислоты глутамата через его ионотропные рецепторы и АХ (ацетилхолин) через N-холинорецепторы открываются быстрые Na+-каналы, что приводит к деполяризации мембраны и в результате клетка переходит в состояние возбуждения. Процесс возбуждения необходим для всех основных функций мозга: психологической и физической активности, целесообразного поведения, обучения, памяти, восприятия чувствительных и болевых импульсов. Наоборот, ГАМК (тормозной медиатор головного мозга) и глицин (тормозной медиатор спинного мозга), взаимодействуя с рецепторами, открывают Сl--каналы, что приводит к гиперполяризации, и в результате возникает торможение клетки. Оно столь же необходимо для всех функций клетки, как и возбуждение, должен существовать баланс возбуждающих и тормозящих нейротрансмиттеров (при бодрствовании преобладают первые, при сне – вторые).
При эпилепсии происходит сдвиг к преобладанию глутамата, что приводит к судорогам. Большинство противоэпилептических лекарств, у которых разные механизмы действия, так или иначе, стимулируют систему ГАМК. Активация синтеза ГАМК (вальпроат) восстанавливает баланс и тоже дает противосудорожный эффект. При нарушении кровоснабжения головного мозга (инсульт) глутамата выделяется много с последующей аккумуляцией ионов Са2+ – важный механизм повреждения и даже гибели нейронов. При попадании в рану возбудителя столбняка он вырабатывает токсин, который выключает рецепторы ГАМК. Она не может работать и тогда возбуждающие нейротрансмиттеры, не встречая противодействия, вызывают перевозбуждение, что приводит к появлению судорог, а иногда и к смерти. Судороги при отравлении стрихнином, блокирующим рецептор глицина, тоже возникают от нарушения баланса в сторону преобладания эффектов возбуждающих медиаторов.
На ГАМКА-рецепторы действуют три группы веществ, которые тормозят головой мозг, увеличивая действие эндогенной ГАМК:
барбитураты – средства для неингаляционного наркоза и снотворные;
бензодиазепины – транквилизаторы (успокаивающие).
нейростероиды, которые синтезируются в нейронах или являются метаболитами прогестерона, гормона желтого тела и плаценты.