

4 КУРС (ЭМБРИОЛОГИЯ) / Несортированное по эмбриологии / ТАБЛИЦА
.pdfV. Гаметогенез |
71 |
трируются в клетках внутренних областей зародыша, поблизости от так называемого бластоцеля. Окончательное обособление первичных гоноцитов происходит в этой группе животных на стадии поздней бластулы, когда зародыш насчитывает несколько сотен клеток, но основные зачатки органов еще не обособились.
Значительно позже происходит обособление первичных гоноцитов у хвостатых амфибий. В противоположность бесхвостым амфибиям этот процесс идет не автономно, а под влиянием других, соседних эмбриональных тканей. Гоноциты хвостатых амфибий возникают из среднего зародышевого листка (мезодермы) под воздействием внутреннего листка (энтодермы). Это воздействие осуществляется на стадии бластулы, само же обособление гоноцитов на стадии гаструлы или даже нейрулы, т. е. в период закладки других зачатков органов.
У птиц, по современным данным, первичные половые клетки возникают на ранних стадиях развития зародыша из наружного листка (эпибласта), а затем мигрируют во внутренний листок (гипобласт), в составе которого располагаются у переднего конца зародыша, в области так называемого головного серпа.
Возможно, что и у млекопитающих первичные половые клетки возникают из наружного листка зародыша. Позже они перемещаются к так называемому устью желточного мешка. У зародыша человека окончательное обособление гоноцитов от соматических клеток происходит к 56-му дню развития; к этому времени сформированы зачатки основных органов.
Половые клетки у круглых червей, членистоногих, позвоночных – первичные гоноциты, обособляющиеся от клеток соматических на ранних стадиях развития. Иными словами, дифференцировка клеток зародыша на половые и соматические сопровождается у данных групп животных также и их необратимой детерминацией.
Однако далеко не у всех групп животных гоноциты обособляются исключительно на ранних стадиях эмбрионального развития и не могут позже пополняться за счет соматических клеток. У низших многоклеточных – губок, стрекающих кишечнополостных, мшанок, а также у некоторых кольчатых червей и полухордовых имеются особые тотипотентные стволовые клетки (способные трансформироваться в любой тип клеток данного
72 |
Болотов А. В. Биология размножения и развития |
вида), которые в течение всей жизни организма пополняют запас его половых клеток. Таким образом, у этих организмов линии половых и соматических клеток практически неотделимы.
У плоских и кольчатых червей стволовые клетки, называемые необластами, присутствуют во многих тканях. Поэтому половые клетки могут у них возникать и в случае регенерации из небольших участков тела взрослых животных при полном удалении половых желез. У продолжительно голодающих планарий (плоские черви) половые клетки могут дедифференцироваться и превращаться в стволовые клетки, используемые для регенерации соматических тканей. Впоследствии из них могут вторично возникать половые клетки. Вместе с тем у многих кольчатых червей имеется и рано обособляющаяся линия эмбриональных гоноцитов. Таким образом, у них существуют два источника половых клеток; раннеэмбриональный и соматический.
Важно напомнить также, что почти у всех растений (за исключением зеленых водорослей) линии половых и соматических клеток фактически неотделимы на протяжении всей жизни: половые клетки выделяются из соматических только при формировании мужских и женских гаметофитов.
Каков бы ни был источник половых клеток – раннеэмбриональные первичные гоноциты или постоянно существующие резервные клетки, они должны пройти длинные пути превращений, прежде чем стать зрелыми, способными к оплодотворению половыми клетками.
2. Состав, развитие и особенности половых клеток
Половые, или зародышевые, клетки (гаметы) представляют собой особые клеточные протоплазматические образования, из которых после их слияния во время оплодотворения начинает развиваться новый индивидуум. Половые клетки делятся на мужские (спермии, сперматозоиды) и на женские (яйцеклетки, овули). При оплодотворении обе половые клетки сливаются в одно общее клеточное целое (оплодотворенное яйцо, spermovium, или зигота). Процесс оплодотворения одновременно служит для зиготы импульсом к интенсивному клеточному делению
V. Гаметогенез |
73 |
(сегментации, или дробящему делению), которое является первым шагом в развитии нового организма.
В соответствии с основным биогенетическим законом в живом веществе половых клеток сконцентрирована совокупность свойств, являющаяся результатом длительного процесса филогенетического развития соответствующего вида животного и всего животного мира в целом. Таким образом, эта совокупность возникает как результат влияний внутренней и внешней среды на организмы во время их филогенеза и представляет собой основу наследственности будущей особи, зафиксированную, прежде всего в ядерном веществе половых клеток.
Половые клетки, из которых развивается новый организм, являются биологически наиболее сложными клетками; они характеризуются максимальными возможностями развития, в то время как все остальные клетки, возникающие из оплодотворенного яйца в процессе развития организма или же при дифференциации отдельных тканей, являются биологически менее сложными и имеют меньшие возможности развития (меньший проспективный потенциал). В связи с этим половые клетки можно считать завершением цикла развития организма, его концентрированием, а в филогенетическом отношении одновременно началом развития новых организмов. В половых клетках как бы кумулируется весь путь развития, который прошли организмы предыдущих поколений.
Особенности половых клеток
1.Измененное ядерно-плазменное отношение по сравнению
ссоматическими клетками;
2.Гаплоидный наследственный материал (nc).
3.Измененный обмен веществ (половые клетки находятся как бы в состоянии депрессии).
4.Наличие специфических структур: акросомальный аппарат, хвостик для движения спермия, оболочки яйца, наличие кортикальных и желточных гранул.
5.Отсутствие ряда органелл в клетках: в сперматозоидах (аппарат Гольджи, эндоплазматический ретикулум, рибосомы) и яйцеклетках (центриоль).
6.Незаконченный жизненный цикл.
74 |
Болотов А. В. Биология размножения и развития |
3. Строение половых клеток
Сперматозоид
Спермий, сперматозоид (мужская половая клетка) – зрелая мужская половая клетка – по своим морфологическим и физиологическим свойствам значительно отличается от женской половой клетки – яйца.
Спермий был открыт в 1677 г. учеником Левенгука Л. Гаммом. Сам Левенгук считал спермий (бойко движущиеся образования с характерным хвостиком) примитивными зародышами, паразитирующими в семени и называл их «семенными зверьками». От него происходит и более старое, до сих пор иногда применяемое название мужской половой клетки – сперматозоон (от лат. animalculum – семенное животное). В соответствии с господствовавшими в то время представлениями, головка этого семенного животного содержала в миниатюре всю будущую особь. Термин «сперматозоид» ввёл в начале XIX в. Карл Эрнст фон Бэр.
По своему виду спермий значительно отличается не только от яйцеклетки, но и от всех остальных клеток организма. В отличие от крупной и инертной яйцеклетки сперматозоиды значительно более мелкие, размер которых у разных видов животных, колеблется от 20 мкм (у крокодила), до 2 мм в длину (у амфибии); они активны и подвижны.
Ядерно-плазменное отношение сильно смещено в сторону ядра, т. е. у них почти нет цитоплазмы. За счет уникального механизма конденсации хроматина (из ядра удаляются гистоны, и ДНК связывается с белками-протаминами), большая часть цитоплазмы выбрасывается из сперматозоида в виде так называемой «цитоплазматической капли», остаются только самые необходимые органеллы. Поэтому они утрачивают и некоторые органеллы: рибосомы, эндоплазматический ретикулум, аппарат Гольджи. В ядре сперматозоида человека содержится 23 хромосомы, одна из которых является половой (X или Y), остальные – аутосомами. Среди спермиев 50 % содержат Х-хромосому и 50 % – Y-хромосому. Показано, что масса Х-хромосомы больше массы Y-хромосомы, поэтому спермии, содержащие Х-хромосому, менее подвижны, чем содержащие Y-хромосому. Сперматозоиды,
V. Гаметогенез |
75 |
содержащие Y-хромосому, называются андроспермиями, Х-хромосому – гиноспермиями. В спермограмме здорового мужчины наряду с нормальными встречаются и патологические формы спермиев, но обычно не более 20–25 % (рис. 14, Б).
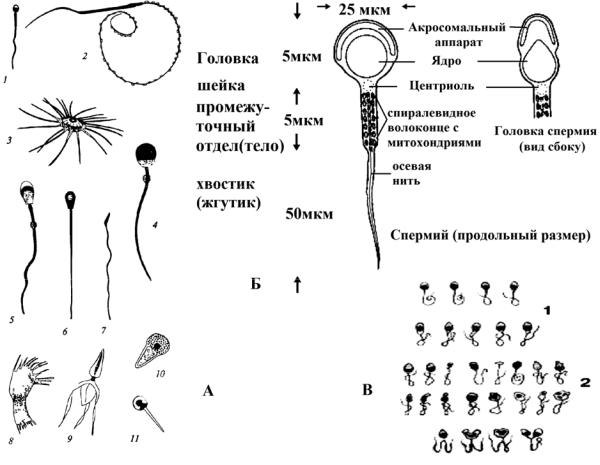
Рис. 14. Разные виды сперматозоидов (А):
1 – человек; 2 – тритон; 3 – речной рак; 4 – морская свинка; 5 – хряк; 6 – бык; 7 – петух; 8 – ветвистоусый рак; 9 – десятиногий рак; 10 – лошадиная аскарида; 11 – острица.
Схема строения зрелого сперматозоида человека (Б). Сперматозоиды человека нормальные (1) и аномальные (2) (В)
Форма сперматозоидов очень разнообразна, но среди них можно выделить 2 главных типа: жгутиковые и безжгутиковые. Безжгутиковые формы сравнительно редки (круглые черви, высшие раки, моллюски), которые имеют причудливую форму
(рис. 14, А).
В жгутиковых сперматозоидах, как правило, присутствует головка, тело, шейка и хвостик (рис. 14, А). У сперматозоида человека, например, различают четыре основные части (рис. 14, Б):
76 |
Болотов А. В. Биология размножения и развития |
1.Головка сперматозоида человека имеет форму эллипсоида, сжатого с боков, с одной из сторон имеется небольшая ямка, поэтому иногда говорят о «ложковидной» форме головки сперматозоида у человека (рис. 14, Б). Передняя часть головки покрыта тонким, прозрачным слоем плазмы, которая является довольно устойчивой и умеренно заостренной. У некоторых животных это приспособление, возможно, играет роль образования, облегчающего проникновение головки спермия в яйцеклетку во время, оплодотворения.
В передней части головки расположен акросомальный аппарат (рис. 14, Б), состоящей из секреторного пузырька – акросомы (содержащий гидролитические ферменты, и позволяющий спермию проникнуть через наружные яйцевые оболочки). Акросома функционально соответствует лизосоме. Она развивается из аппарата Гольджи и содержит около 15 гидролитических ферментов – гиалуронидазу и несколько протеиназ, основным из который является трипсиноподобный акрозин – они осуществляют «переваривание» гранулярного слоя и плазматической мембраны ооцита непосредственно перед оплодотворением. Акросома лежит спереди от ядра и покрывает собой половину ядра (поэтому часто акросому сравнивают с шапочкой). При контакте
сяйцеклеткой акросома выбрасывает свои ферменты наружу и растворяет небольшой участок оболочки яйцеклетки, благодаря чему образуется небольшой «проход» для проникновения сперматозоида.
Почти все вещество головки – за исключением очень тонкого и почти невидимого цитоплазматического слоя на поверхности – состоит из ядерного вещества, очень сгущенного в этой части и интенсивно красящегося ядерными красками. В связи с сильной конденсацией хроматин неактивен – в ядре сперматозоида не синтезируется РНК. Иногда в ядерном веществе головки обнаруживается шаровидная вакуоль. Длина головки составляет приблизительно 4–5 мкм.
2.Шейка спермия (рис. 14, Б) представляет собой короткую, более узкую часть, образованную мягким, гомогенным плазматическим веществом. Сразу же за головкой в ней располагается проксимальная (передняя) центриоль, по мнению одних авторов,
ввиде двух или нескольких зернышек, по мнению других – в
V. Гаметогенез |
77 |
виде пластинки. Мягкое вещество шейки обусловливает сравнительную подвижность головки по отношению к хвосту и ее наклон под определенным, почти прямым углом. Длина шейки составляет приблизительно 0,5 мкм.
3.Соединяющая (средняя) (рис. 14, Б) часть спермия или тело отграничивается от шейки проксимальной частью дистальной (задней) центриоли в виде пластинки, которая, по мнению некоторых авторов, состоит из целого ряда мелких зернышек. Соединяющая часть оканчивается кольцеподобным образованием, исходящим из дистальной части дистальной центриоли. Посередине соединяющей части проходит осевая нить хвоста (рис. 14, Б), образованная тонкими фибриллами и исходящая, по мнению одних авторов, из передней центриоли, по мнению других – из задних зернышек. Осевая нить выходит из соединяющей части через кольцеподобное образование и проходит далее по хвосту спермия. В области соединяющей части она непосредственно покрыта цитоплазматическим слоем, вокруг которого несколько раз обвивается спиралевидное волоконце, образованное митохондриями цитоплазматического слоя (спираль) (рис. 14, Б). Длина соединяющей части составляет приблизительно 5–6 мкм; она считается двигательным центром хвоста.
4.Хвост, или жгутик (рис. 14, Б), представляет собой собственно продолжение осевой нити соединяющей, средней части. Осевая нить на отрезке длиной в среднем в 20–30 мкм покрыта тонким слоем цитоплазмы, а ее конец, длиной приблизительно в 5 мкм, обнажен и, постепенно утончаясь, заканчивается заострением. Таким образом, общая длина спермия колеблется от 50 до
60 мкм.
Яйцеклетка (ovulum, или женская половая клетка)
Зрелая яйцеклетка относится к наиболее крупным клеткам организма. Следовательно, яйцеклетка поставляет большую часть строительного материала, необходимого для развития нового организма.
Яйцевые клетки разных видов животных и птиц сильно варьируют по размерам, при этом форма у них может быть либо шаровидная, либо овальная. Самое крупное яйцо обнаружено у китовой акулы, длина 60 см, а диаметр – 40 см. Достаточно крупные яйца у тигрового питона – 12 см в длину и с диаметром
78 |
Болотов А. В. Биология размножения и развития |
6 см. Среди птиц самое крупное яйцо у страуса в длину 15–20 см, диаметром 15–18 см. Среди млекопитающих самые крупные яйца свойственны яйцекладущим – утконосу и ехидне. Диаметр яйца утконоса – 4,4 мм, ехидны – 3 мм. Зрелая яйцеклетка человека имеет примерно 150 мкм (0,15 мм) в диаметре, макака – резуса – 118 мкм, морской свинки – 76 мкм, кролика – 160 мкм, а мыши – 80 мкм. А самая мелкая яйцеклетка обнаружена у примитивного млекопитающего – полевки пашенной, размер которой составляет всего 40 мкм.
Яйцевая клетка человека имеет правильную шаровидную форму. Величина живой яйцеклетки человека колеблется от 130 до 160 мкм, но несмотря на свои сравнительно большие размеры, она была открыта довольно поздно.
Хотя о «яичке» упоминалось уже в литературе XVII в., однако в то время яйцеклеткой считали шаровидные, пузырчатые образования, хорошо видимые простым глазом в яичниках животных и человека и известные в настоящее время как овариальные фолликулы (Граафовы фолликулы – Регнерус де Грааф, 1677). Сама яйцевая клетка была открыта лишь в 1827 г. Карлом Бэром. Однако уже и другие авторы – современники Бэра – в первой половине XIX в. под различными названиями описывали некоторые самостоятельные составные части яйцеклетки. Так, Ян Пуркинье обнаружил ядро птичьей яйцеклетки и назвал его vesicula germinativa (1825), Рудольф Вагнер (1836) описал в нем ядрышко. Большую часть шаровидного клеточного тела яйцеклетки составляет цитоплазма, которая в данном случае называется ооплазмой. В живой яйцеклетке ооплазма представляет собой мягкое, прозрачное, зернистое вещество; более отчетливо зернистость видна в центральных отделах клетки, в области, расположенной вокруг ядра, в то время как более поверхностный слой ооплазмы или вообще не содержит никаких гранул, или же содержит их в малом количестве. Некоторые зернышки в ооплазме соответствуют зернистым митохондриям, которые обнаруживаются в яйцевой клетке при помощи соответствующих гистологических методов. Однако большую часть гранул составляет желтковое вещество. Объем яйца, как правило, превосходит объем спермия во много раз (у морского ежа их отношение составляет 10 000:1). Ядерные гаплоидные компоненты
V. Гаметогенез |
79 |
у них примерно равны, а вот по содержанию цитоплазматических компонентов различия велики. Цитоплазма яйца представляет собой огромную кладовую запасов, накопленных в период вителлогенеза. Эти запасы включают белки, рибосомы, транспортную и матричную РНК, морфогенетические факторы. Это молекулы, которые направляют процессы дифференцировки клеток, необходимые для образования различных тканей и органов. Они рассеяны по всему яйцу и распределяются между разными клетками при дроблении.
|
Важной отличительной спо- |
|
|
собностью яйцеклеток является |
|
|
то, что ядерно-плазменное отно- |
|
|
шение смещено в сторону цито- |
|
|
плазмы (рис. 15). |
|
|
Ядро яйцевой |
клетки – ша- |
|
ровидное, располагающееся чаще |
|
|
всего несколько |
эксцентрично |
|
(рис. 16, 1). В ядре яйцеклетки |
|
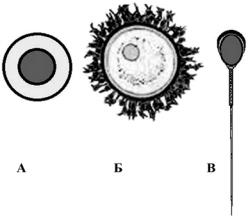
Рис. 15. Соотношение |
человека содержится 23 хромо- |
|
ядерного вещества к цито- |
сомы; одна из них является поло- |
|
плазму у соматический (А) |
вой Х-хромосомой. |
|
клетки, яйцеклетки (Б) и |
На фиксированных и окра- |
|
сперматозоида (В) |
шенных препаратах видно, что в |
|
ядре содержится мало хроматина, в связи с чем оно слабо окрашивается и имеет везикулярный (пузырчатый) вид. Хроматин носит волокнистый характер и в периферических отделах ядра образует более компактный слой, представляющий собой нечто наподобие ядерной оболочки. В ядре, как правило, находится крупное шаровидное псевдоядрышко.
В зрелой яйцеклетке центриоль уже не содержится, поскольку она исчезает в процессе развития и созревания яйца.
Все компоненты яйца располагаются в цитоплазме яйца ассиметрично, обеспечивая полярность яйца и эмбриона в будущем (рис. 16). То есть яйцо имеет два полюса: анимальный и вегетативный. Та часть яйца, ближе к которой располагается ядро или на котором выделяются редукционные тельца называется анимальным, а противоположный – вегетативным. А ось, которая проходит через эти два полюса называется анимально-
80 |
Болотов А. В. Биология размножения и развития |
вегетативной осью клетки, играющей большое значение в процессах дробления и дальнейшего развития организма. Эта ани- мально-вегетативная поляризация яйцеклетки решающим образом ориентирует последующие морфогенетические процессы: за редкими исключениями первые две борозды делений дробления оплодотворенной яйцеклетки проходят по взаимно перпендикулярным анимально-вегетативным меридианам, пересекаясь на аномальном и вегетативном полюсах, а у взрослых животных передне-задняя ось тела либо совпадает с анимальновегетативной осью яйцеклетки (так называемые протаксонные животные – многощетинковые черви, позвоночные и др.), либо перпендикулярна ей (плагиаксонные животные – малощетинковые черви, некоторые членистоногие).
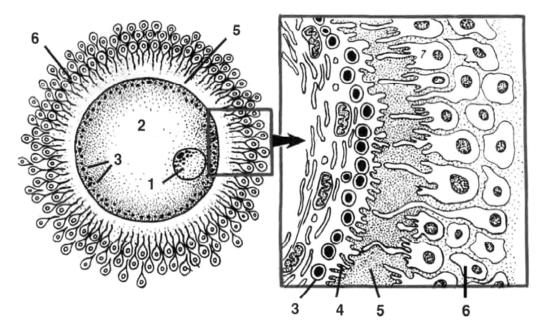
Рис. 16. Схема строения яйцеклетки:
1 – ядро; 2 – цитоплазма с желточными гранулами; 3 – кортикальные гранулы; 4 – микроворсинки на поверхности плазмолеммы; 5 – блестящая оболочка; 6 – зернистая оболочка.
Первые морфологические проявления поляризации яйцеклетки приурочены к периоду вителлогенеза: у большинства яйцеклеток желток откладывается преимущественно в вегетативном полушарии, а ядро ооцита оттесняется в анимальное полушарие, где больше свободной цитоплазмы. Но только в период делений созревания, а точнее, в период выделения 2-го ре-