

4 КУРС (ЭМБРИОЛОГИЯ) / Несортированное по эмбриологии / ТАБЛИЦА
.pdfVII. Дробление |
141 |
червей, плоских червей и моллюсков, то оказалось, что у клеток, занимающих одно и то же положение, судьба одинакова.
В-четвертых, первые три деления дробления происходят синхронно.
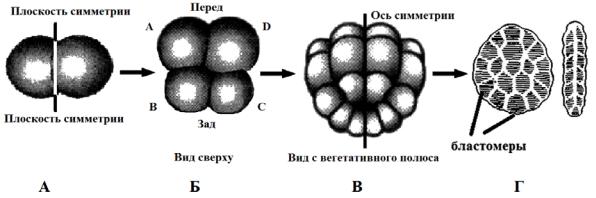
Первые два дробления почти меридиональны и приводят к образованию 4 крупных клеток – макромеров (A, B, C, D) (рис. 30, А). У многих видов эти бластомеры различаются по размерам, при этом самыми крупными обычно бывают D. При последующих делениях каждый макромер на своем анимальном полюсе отделяет маленький микромер (рис. 30, Б).
Каждый последовательно появляющийся квартет микромеров смещается вправо или влево относительно сестринских макромеров, это и приводит к характерному расположению бластомеров по спирали.
Возникающие впоследствии бластулы не имеют бластоцеля и носят название стерробластула (рис. 30, В).
3. Билатеральное голобластическое дробление
Характерен для нематод, а также для многих низших хордовых, в том числе для асцидий, аппендикулярий. Характерной особенностью этого типа является раннее проявление билатеральной симметрии.
Замечательная особенность этого типа дробления заключается в том, что плоскость первого деления устанавливает единственную плоскость симметрии зародыша (рис. 31, А). Каждое последующее деление ориентируется по отношении к этой плоскости симметрии так, что половина зародыша по одну сторону от первой борозды представляет зеркальное отражение половины зародыша по другую сторону. Второе деление, так же как и первое, – меридиональное, но в отличие от первого борозды дробления не проходят через центр яйца. В результате возникают два крупных передних бластомера (A, D) и два задних меньшего размера (рис. 31, Б). На каждой стороне теперь имеются один крупный и один небольшой бластомер. При последующих делениях различия в размерах и форме бластомеров только подчеркивают билатеральную симметрию зародыша (рис. 31, В).
142 |
Болотов А. В. Биология размножения и развития |
Рис. 31. Билатеральное голобластическое дробление (А, Б, В) и образование плакулы (бластула) (Г)
На 32 клеточной стадии формируется бластула с небольшим бластоцелем и начинается гаструляция. Бластула у асцидий называется плакула (новолат. placula, от греч. plax – плоскость, пластинка), которая имеет вид двуслойной пластинки, образованной более или менее однородными клетками, между слоями которой есть небольшая полость (рис. 31, Г).
4. Асинхронное, ротационное голобластическое дробление
Эмбрионы млекопитающих развиваются, как правило, внутриутробно. По этой причине яйцеклетка у них олиголецитальная (мало желтка) и изолецитальная (желток равномерно распределен).
После оплодотворения зигота постепенно спускается по маточной трубе в полость матки. В этот период, в течение примерно трех дней, зигота проходит стадию клеточного деления, известную как дробление. При дроблении число клеток увеличивается, но общий их объем не меняется, так как каждая дочерняя клетка мельче, чем исходная. Дробление полное, но неравномерное и асинхронное.
Дробление у млекопитающих отличается от всех других типов дробления по многим признакам:
1.Первая особенность заключается в относительно медленном темпе делений.
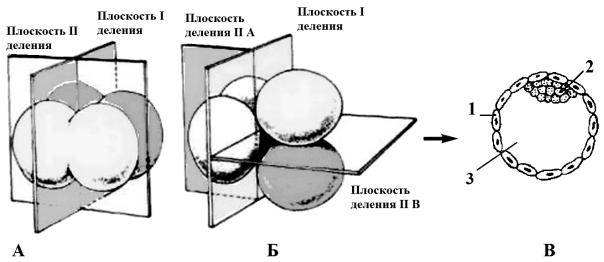
2.Своеобразное расположение бластомеров относительно друг друга. Первое деление – меридиональное. В ходе второго
VII. Дробление |
143 |
деления один бластомер делится меридионально, но перпендикулярно первому делению, а второй – экваториально. Поэтому такой тип дробления был назван чередующимся или ротационным (рис. 32, Б).
Рис. 32. Сравнение радиального (А) и асинхронного, ротационного дробления млекопитающих (Б) и образование бластоцисты у млекопитающих (В).
А – первое и второе деление радиального дробления; Б – первое и второе деление асинхронного, чередующегося дробления. В – образование бластоцисты: 1 – трофобласт; 2 – зародышевый узелок (эмбриобласт); 3 – бластоцель
3.Выраженная асинхронность раннего дробления. Бластомеры у млекопитающих не делятся все одновременно, поэтому у зародышей не происходит равномерного нарастания числа бластомеров. Число которых может быть нечетным.
4.Явление компактизации. Бластомеры на 8-й клеточной стадии расположены рыхло. Однако после третьего деления поведение бластомеров резко изменяется. Они внезапно сближаются и площадь контакта между клетками максимально увеличивается, и они образуют плотный клеточный шар – морулу. Эта компактная упаковка стабилизируется плотными контактами, которые образуются между клетками, расположенными на поверхности шара, и изолируют лежащие внутри клетки. Между клетками внутри шара образуются щелевые контакты, через которые могут молекулы и ионы диффундировать из клетки в клетку.
144 |
Болотов А. В. Биология размножения и развития |
5. Образование бластулы, которая называется бластоциста (рис. 32, В). Первоначально морула не имеет внутренней полости, но в процессе кавитации наружные клетки, которые называются трофобластом секретируют в морулу жидкость, что и приводит к образованию полости бластулы. А внутренние клетки, которые называются внутренней клеточной массой располагается на одной стороне стенки полого шара.
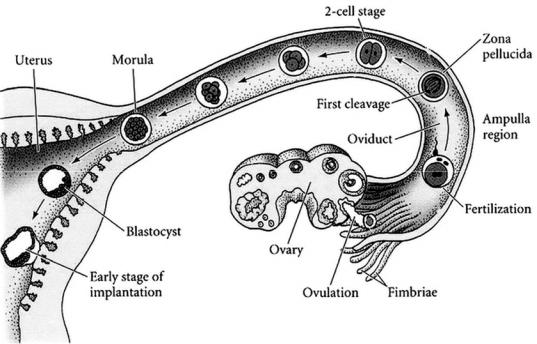
В течение первых суток оно происходит медленно. Первое деление завершается через 30 ч. За стадией двух бластомеров следует стадия трех бластомеров. Второе дробление наступает через 10 ч после первого и приводит к образованию четырехклеточной стадии. Через 40 ч образуются 4 клетки. Дробящийся зародыш постепенно перемещается по яйцеводу и в нижней его части достигает восьмиклеточной стадии (рис. 33). Примерно через 50–60 ч после оплодотворения достигается стадия морулы – шара из 16 и более клеток.
Затем один из них делится, а второй по времени запаздывает с делением. Клетки бластомеры, которые делятся чаще – бледные и мелкие, а те, которые делятся реже – крупные и темные.
Рис. 33. Схема развития зародышей млекопитающих от оплодотворения до имплантации (по Гилберту С., 1993)
VII. Дробление |
145 |
Клетки эмбриона, бластомеры, укладываются компактнее (происходит компактизация), дифференцируются на внутренние и наружные, различающиеся строением и будущей судьбой. Следующие деления приводят к образованию морулы в виде плотного комка клеток, на 3–4-е сутки начинается формирование бластоцисты, напоминающую полый пузырек, заполненный жидкостью. Ее трофобласт – тонкая стенка пузырька – образован вытянутыми крупными клетками, потомками наружных клеток морулы, а внутри находится небольшое скопление мелких клеток, асимметрично прикрепленных к стенке. Это – внутрен-
няя клеточная масса (ВКМ), или эмбриобласт. На стадии бла-
стоцисты зародыш попадает в матку, выходит из оболочки оплодотворения (zona pellucida) и прикрепляется к стенке матки с помощью трофобласта, который затем станет эмбриональной частью плаценты.
5. Дискоидальное меробластическое дробление
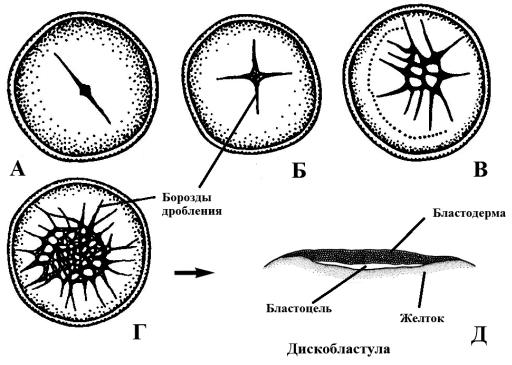
Дробление резко телолецитальных яйцеклеток (рептилии, птицы, костистые рыбы) называется дискоидальным. Так происходит дробление у костистых рыб (рис. 34).
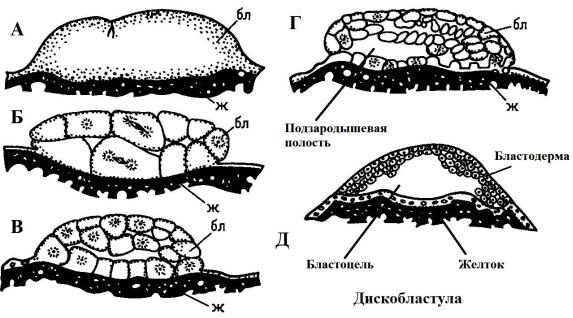
Рис. 34. Дробление яйца у костистых рыб (А, Б, В, Г) и образование бластулы (Д),
бл – бластомеры; ж – желток
146 |
Болотов А. В. Биология размножения и развития |
Вскоре после оплодотворения яйцеклетки основная масса цитоплазмы, которая ранее тонким равномерным слоем накрывала желток, перетекает к анимальному полюсу, где с самого начала было расположено ядро зиготы. Поскольку деления дробления не распространяются на богатую желтком цитоплазму, на ранних стадиях дробления клетки у своего основания не отделены друг от друга, т. е. не образуются отдельные клетки (рис. 34, А). В результате на анимальном полюсе тангенциальными делениями, которые проходят параллельно поверхности зиготы, образуется однослойная бластодерма (рис. 34, В, Г). Затем дробления идут перпендикулярно, так образуется несколько слоев клеток зародыша. Таким образом формируется сначала однослойная, а затем и многослойная группа бластомеров – дискобластула. Между бластодермой и желтком имеется пространство, называемое подзародышевой полостьюилибластоцелью (рис. 34, Д).
Дискоидальное дробление птиц (рис. 35) идет по сходному пути, с той главной разницей, что бластодиск, лежащий на желтке, значительно тоньше, нежели у костистых рыб, и поэтому должно пройти больше делений дробления, прежде чем дискоб ластула станет многослойной. К тому времени, когда птица
Рис. 35. Дробление яйца у птиц (А, Б, В, Г) и образование бластулы (Д)
VII. Дробление |
147 |
откладывает яйцо, бластодерма содержит около 60 тысяч клеток и клетки начинают выселяться в подзародышевую полость, где образуется второй слой. Таким образом зародыш птицы имеет 2 слоя клеток: наружный – эпибласт и лежащий под ним гипобласт. Таким образом, в результате этого типа дробления образуется дискобластула (рис. 35, Д). Она включает три компонента:
1)плоский зародышевый диск, образующийся из апикальной части зиготы и состоящий из небольших клеток;
2)нераздробившийся желток;
3)бластоцель – узкая щель между диском и желтком.
6.Поверхностное меробластическое дробление
Центролецитальным яйцам насекомых, очень богатых желтком, свойственен этот тип дробления. Из-за большого количества желтка, расположенного центрально, дробление ограничено только поверхностным слоем цитоплазмы.
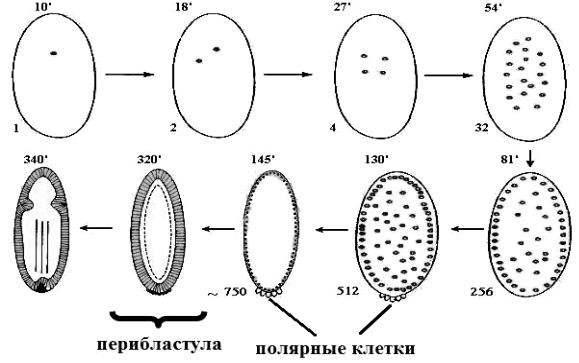
Рис. 36. Поверхностное дробление зародыша дрозофилы.
256 ядер – стадия сенцитиальной бластодермы; 512 ядер – формирование полярных клеток, начало обособления клеток бластодермы; 750 ядер – формирование клеток бластодермы; 320´ – стадия клеточной бластодермы, бластуляция; 340´ – начало формирования зародышевой полоски. Цифры над зародышем соответствуют числу минут; цифры внизу обозначают число ядер
148 |
Болотов А. В. Биология размножения и развития |
Одна из примечательных особенностей этого типа дробления заключается в том, что обособление клеток происходит только после многократного разделения ядер. На стадии 256 ядер, ядра с островками цитоплазмы (энергиды) мигрируют на периферию яйца, примерно на 81-й минуте, где продолжаются деления, но с убывающей скоростью. Теперь зародыш носит название синцитиальной бластодермы. После образования полярных клеток плазматическая мембрана яйца образует выпячивания, которые углубляются между ядрами и в конечном счете обособляется каждое из них в отдельной клетке. Так возникает клеточная бластодерма, спустя почти 5 часов, клетками которой однослойным покровом одевают богатую желтком сердцевину яйца. Так образуется бластула, называется перибластула, пузырёк, стенка которого состоит из одного слоя клеток, а полость заполнена нераздробившимся желтком
(см. рис. 36).
VIII. ГАСТРУЛЯЦИЯ
Несмотря на большое разнообразие процессов гаструляции, можно дать вполне определенную классификацию гаструл и путей возникновения их из предыдущей стадии бластулы. К сожалению, еще не создана классификация, которая исходила бы из принципа эволюции, так как историческое происхождение разных типов гаструляции изучено недостаточно, и эмбриология имеет в этой области больше научных загадок, чем достоверных сведений.
Некоторые способы гаструляции встречаются во всех типах животного мира.
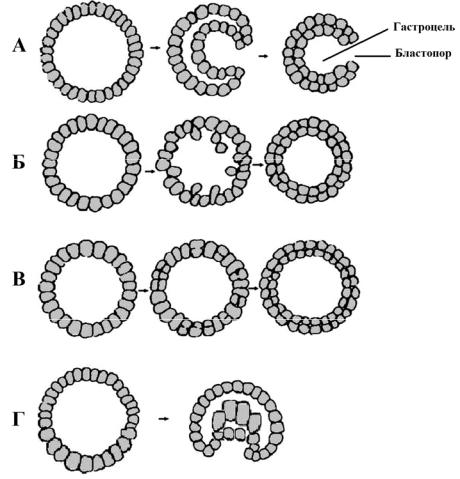
1. Инвагинационная гаструла (рис. 37, А) образуется пу-
тем инвагинации (впячивания) части однослойной бластодермы (стенки бластулы) внутрь бластоцеля. Грубое подобие такого типа гаструляции – результат впячивания стенки резинового мячика при надавливании на него. Впячивание можно довести вплотную до внутренней стенки мячика (на противоположном от места вдавливания участке), или вдавливание можно сделать не столь значительным. Образуется двухслойный мешок, наружной стенкой которого является первичная эктодерма, а внутренней – первичная энтодерма. Впячивание образует архентерон (первичный кишечник), а отверстие, при помощи которого он сообщается с наружной средой, называется первичным ртом, или
бластопором (рис. 37, А).
Судьба бластопора у разных животных неодинакова. У многих животных первичный рот, соответственно развиваясь и дифференцируясь, превращается в дефинитивный рот взрослого организма. Эти животные называются первичноротыми (Protostomia). К первичноротым относятся черви, моллюски и членистоногие.
Не менее обширна группа вторичноротых (Deuterostomia), у которых бластопор превращается или в анальное отверстие, или (у хордовых) в нервно-кишечный канал, находящийся в заднем конце эмбриона. Рот у таких животных возникает на переднем конце, на брюшной стенке, в результате особых формообразовательных процессов (впячивание эктодермы, прободение стенки
150 |
Болотов А. В. Биология размножения и развития |
средней кишки). К Deuterostomia относятся: щетинкочелюстные, плеченогие, иглокожие, кишечнодышащие и хордовые.
Рис. 37. Разные способы гаструляции.
A – инвагинационная гаструла; Б – иммиграционная гаструла; В – деляминационная гаструла; Г– эпиболическая гаструла
Указанными признаками отнюдь не исчерпывается необходимость разделения животных на две группы: первично- и вторичноротые. В дальнейшем будут сообщены иные не менее важные признаки обеих групп.
Очень многие вопросы являются спорными потому, что биология еще не знает истинных предков современных; нет единого мнения и о филогенетическом значении разного типа гаструл. Многие современные исследователи, присоединяясь к И. И. Мечникову, считают исходной формой гаструлы иммиграционную гаструлу и выводят все способы образования энтодермы из мультиполярной иммиграции и деляминации.