

4 КУРС (ЭМБРИОЛОГИЯ) / Несортированное по эмбриологии / ТАБЛИЦА
.pdfVIII. Гаструляция |
151 |
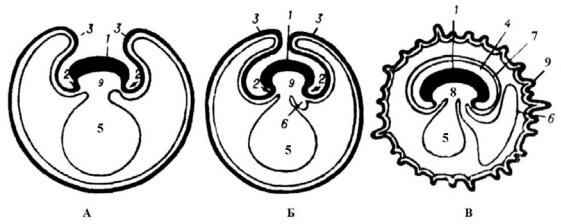
Возвращаясь к характеристике инвагинационной гаструлы, отметим, что в ней имеются две полости: в большей или меньшей степени сохраняется бластоцель (хотя в некоторых случаях эта полость почти нацело вытесняется впячиванием) и возникает новая полость – гастроцель (рис. 37, А), или гастральная полость или полость первичного кишечника (архентерон).
2. Иммиграционная гаструла (рис. 37, Б) характерна для многих кишечнополостных. Она развивается вследствие активного выселения части клеток стенки бластулы внутрь бластоцеля. Это выселение клеток в случаях униполярной иммиграции происходит лишь с одного полюса (вегетативного). Выселившиеся клетки образуют позднее внутренний слой гаструлы (энтодерму). Таким образом, как и при инвагинации, зародыш делается двухслойным. Так происходит гаструляция, например, у гидроидного полипа Glytia, гидромедузы Aequorea или у медузы Octorchis gegenbauri. Униполярная иммиграция свойственна почти всем гидромедузам, особенно тем, для которых характерна плавающая целобластула.
Клетки бластодермы могут проникать в бластоцель не в ка- ком-либо одном участке, а по всей поверхности зародыша, причем такая иммиграция может начинаться уже на стадии 16 бластомеров. Эта так называемая мультиполярная иммиграция свойственна, например, зародышу медузы Solmundella, что обнаружил впервые И. И. Мечников. Мультиполярная иммиграция, однако, редкое явление.
У многих видов кишечнополостных, которым свойственна иммиграционная гаструла, происходит столь массовое активное выселение клеток бластодермы, что бластоцель полностью исчезает, так как заполняется плотной массой клеток. Как видим, у иммиграционной гаструлы нет бластопора и, значит, нет характерного для инвагинационной гаструлы сообщения гастроцеля с внешней средой. Можно лишь, проводя далеко идущую аналогию (оправданную представлением об общности животного мира), говорить о месте бластодермы, соответствующем бластопору, что, однако, доступно критике. В случае мультиполярной иммиграции еще меньше оснований говорить о бластопоре или даже указывать какой-либо соответствующий ему участок бластодермы.
152 |
Болотов А. В. Биология размножения и развития |
3.Деляминационная гаструла (рис. 37, В), встречающаяся также у кишечнополостных, впервые описана И. И. Мечниковым. Морфогенетические процессы, сопровождающие этот редкий случай гаструляции, очень своеобразны и в типичном случае кажутся простыми. Яйца некоторых сцифомедуз, например Geryonidae, имеют различимые, концентрически расположенные участки цитоплазмы: один участок – плотная, зернистая эктоплазма и другой – ячеистого строения эндоплазма, в который заходит наружный эктоплазматический слой, содержащий полужидкий желток. Структура яйца такова, что оно может быть отнесено к центролецитальным яйцам.
У таких яиц происходит относительно равномерное синхронное дробление, так что образуется зародыш, состоящий из 2, 4, 8, 16 и, наконец, 32 бластомеров, кажущихся одинаковыми по размерам и пространственному положению.
Дальнейшие дробления бластомеров происходят параллельно поверхности зародыша, и в результате дроблений наружный слой бластомеров будет состоять из одной (бывшей) эктоплазмы, а внутренний – из эндоплазмы и частично из эктоплазмы. Происходит деляминация (расщепление) одного слоя клеток на два. В дальнейшем дробятся только «внутренние» бластомеры, притом опять-таки параллельно поверхности зародыша; возникают, таким образом, две легко различимые клетки: наружный, «эктоплазматический» бластомер и внутренний, «эндоплазматический». В результате такой своеобразной гаструляции зародыш оказывается шаром, состоящим из 64 плоских клеток, образующих наружный слой – эктодерму, и из 32 более высоких клеток внутреннего слоя – энтодермы.
У животных, для которых характерна типичная морула (например, у полипа Clava squaniata), процесс гаструляции происходит так, что клетки, находящиеся на периферии, приобретают отличное от других клеток строение. Пласт первых клеток образует эктодерму, а внутренний пласт – энтодерму. Такой тип гаструляции можно рассматривать как один из вариантов деляминации.
4.Эпиболическая гаструла (рис. 37, Г) формируется у не-
которых животных с резко выраженным телолецитальным строением яиц, например у Bonellia. Значительного размера медленно делящиеся бластомеры (макромеры), с большим содержанием желтка, не обнаруживают способности к каким-либо
VIII. Гаструляция |
153 |
перемещениям. Их обрастают (наползают на них) более быстро делящиеся мелкие, лежащие на поверхности клетки (микромеры). Выходит, что у зародышей таких животных бластопор отсутствует, архентерон не формируется. Только впоследствии, когда макромеры, делясь, становятся меньшего размера, образуется (по неясным биохимическим и физическим причинам) полость и формируется зачаток первичного кишечника.
5. Смешанный тип гаструляции. Типы гаструляции ка-
жутся настолько различными, что их невозможно сравнивать. Несомненно, каждый тип гаструляции требует причинного анализа, но все же в некотором отношении разные способы гаструляции сравнимы. Так, своеобразный процесс эпиболии можно рассматривать как один из особенных вариантов инвагинации. Процессы деляминации можно уподоблять процессам иммиграции, конечно, с большими оговорками. Важно отметить далее, что гаструляция у многих животных происходит так, что имеют место и явления инвагинации, и явления эпиболии, и другие морфогенетические процессы. Такова, например, гаструляция у амфибий, пресмыкающихся, птиц, млекопитающих.
Образование мезодермы
У всех животных, за исключением кишечнополостных, в связи с гаструляцией (параллельно с ней или на следующем этапе, обусловленном гаструляцией) возникает и третий зародышевый пласт – мезодерма. Это совокупность клеточных элементов, залегающих между эктодермой и энтодермой, т. е. в бластоцеле. Таким образом, зародыш становится не двухслойным, а трехслойным. У высших позвоночных трехслойное строение зародышей возникает уже в процессе гаструляции, тогда как у низших хордовых и у всех других типов в результате собственно гаструляции образуется двухслойный зародыш.
Вопросы о путях образования мезодермы у разных животных издавна интересовали и сравнительных анатомов, и эмбриологов. Если отвлечься от всех разнообразных деталей формирования мезодермы у разных животных, можно установить два принципиально разных пути возникновения ее: телобластический, свойственный Protosiomia, и энтероцельный, характерный для Deuterostomia.
154 |
Болотов А. В. Биология размножения и развития |
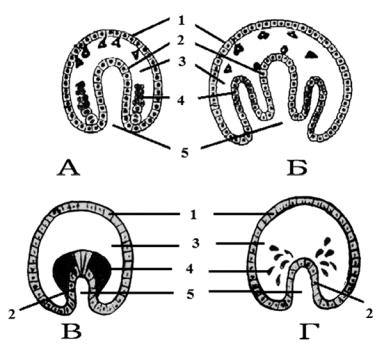
Рис. 38. Образование мезодермы.
А – телобластический путь; Б, В, Г – энтероцельные пути образования мезодермы: собственно энтероцельный, деляминационный, полиферационный. 1 – эктодерма; 2 – эпигастральное впячивание с листком энтодермы; 3 – бластоцель; 4 – формирующаяся мезодерма; 5 – гасторцель
У первичноротых во время гаструляции на границе между эктодермой и энтодермой, по бокам бластопора, уже имеются две большие клетки (или несколько таких клеток-телобластов), отделяющие от себя (вследствие делений) мелкие клетки. Таким образом, формируется средний пласт – мезодерма. Телобласты, давая новые и новые поколения клеток мезодермы, остаются на заднем конце зародыша. По этой причине такой способ образования мезодермы и называют телобластическим (от греч. telos – конец) (рис. 38, А).
Энтероцельные способы (рис. 38, Б, В, Г) образования мезодермы
1. Собственно энтероцельный способ (рис. 38, Б). На бо-
ковых стенках гастрального впячивания образуются карманоподобные выступы, которые направлены внутрь бластоцеля. Эти выступы отшнуровываются от кишечника и отделяются от него в виде мешочков. Полость мешочков превращается в целóм, т. е. во вторичную полость тела, целомические мешки могут подразделяться на сегменты.
VIII. Гаструляция |
155 |
2.Деляминационный (рис. 38, В). Морфогенетические процессы, сопровождающие этот случай образования мезодермы очень редок. Встречается у сцифомедуз, например Geryonidae. В период гаструляции цитоплазма поляризуется, т. е. имеет различной плотности цитоплазму боковых стенок внутренней и наружной поверхности гастрального впячивания: зернистая эктоплазма и ячеистая эндоплазма. Затем боковые стенки, критически поляризованные, расщепляются и отделяются в бластоцель, где через некоторое время один клеточный пласт снова расщепляется и внутри образуется вторичная полость тела или целóм.
3.Полиферационный (рис. 38, Г). Этот тип образования мезодермы встречается у кишечнополостных и других животных
вразвитии. В период гаструляции в силу разных физикохимических процессов из боковых стенок гастрального впячивания начинают активно выселяться клетки внутрь бластоцеля. Выселившиеся клетки образуют позднее два слоя, между которыми развивается вторичная полость тела или целóм.
IX. ОСОБЕННОСТИ РАЗВИТИЯ МЛЕКОПИТАЮЩИХ
Предками млекопитающих являлись, вероятно, яйцекладущие рептилии. Наиболее примитивны из современных млекопитающих также яйцекладущие животные. В подклассе первозверей (Prototheria) яйцо утконоса Ornithorhynchus развивается вне материнского организма; у ехидны Echidna яйцо развивается в кожной складке на брюшной стороне тела, но питание от матери зародыш не получает. Яйца этих животных содержат большое количество желтка, дробление частичное, свойственное телолецитальным яйцам. Эмбриональное развитие во всех существенных чертах сходно с развитием рептилий.
В подклассе сумчатых (Metatheria) зародыши также почти не получают питание от матери. У некоторых сумчатых развивается примитивная плацента, функционирующая непродолжительное время. Зародыши у сумчатых, например, рождаются недоразвитыми, очень маленькими (1–2 см) и донашиваются матерью в сумке, где устанавливается связь пищевода зародыша с соском матери: ее молоко впрыскивается в пищевод. Яйца у этих животных содержат небольшое количество желтка.
У высших млекопитающих, в том числе и у человека, яйцо практически не содержит желточных включений и у него нет в достаточном количестве ни органических, ни неорганических веществ даже для начальных стадий развития. Все необходимое для своего развития зародыш получает с самого начала от материнского организма. Яйцевые клетки плацентарных животных микроскопического размера (100–300 мкм в диаметре), алецитального или изолецитального типа. Как происходила эволюция яиц, и каков характер развития млекопитающих по мере прогресса установления связи зародышей с материнским организмом, эмбриология не знает, хотя и высказаны важные предположения. Очень интересна и вполне аргументирована смелая гипотеза зоолога В. А. Фаусека, согласно которой эволюцию установления все более тесных связей зародыша с материнским организмом надо рассматривать в свете явлений паразитизма: у за-
IX. Особенности развития млекопитающих |
157 |
родыша, как у паразита, развиваются приспособления для использования материнского организма в качестве места обитания и как источника пищи.
Для анализа причин характера дробления яиц млекопитающих недостаточно гипотез о дроблении. Казалось бы, раз яйца млекопитающих практически лишены желтка, их дробление должно быть таким же, как у изолецитальных яиц беспозвоночных и низших хордовых. На самом деле у многих высших млекопитающих дробление кажется неправильным, оно несинхронное, зародыш проходит, например, стадию трех, пяти бластомеров и т. д. Если учесть сложность организации млекопитающих, а также большие видовые особенности в дроблении, закономерно встречающиеся у разных видов млекопитающих, то надо признать, что говорить о дроблении яиц млекопитающих как о «неправильном» имеется еще меньше оснований, чем oб «анархическом» дроблении некоторых кишечнополостных.
У большинства млекопитающих в результате дробления возникает морула, но у некоторых видов (например, у слоноземлеройки или хоботкового прыгунчика) уже на четырехбластомерной стадии появляется бластоцель. Эта стадия развития соответствует бластуле других позвоночных животных.
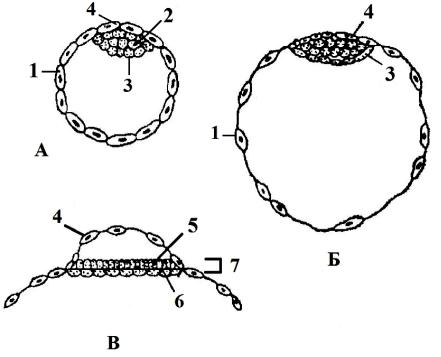
Внутренняя масса бластомеров морулы отличается от поверхностнолежащих своей дальнейшей судьбой. Внутренняя группа бластомеров – эмбриобласт (рис. 39, А, 2) – это источник развития зародыша, его оболочек и желточного мешка, а поверхностно лежащие бластомеры являются трофобластом («питающим зачатком») (рис. 39, А, 1). Эти клетки участвуют в установлении связи зародыша с тканями матки.
Морула млекопитающих превращается в бластоцистубластодермический пузырек, размеры которого у разных животных неодинаковы (у кролика около 7 мм в диаметре). Полость его наполнена жидкостью, вероятно, продуктом жизнедеятельности клеток зародыша. К одному из участков однослойной стенки бластоцисты прилегает скопление клеток эмбриобласта –
зародышевый узелок (рис. 39, 3).
Участок трофобласта, с которым соприкасается эмбриобласт, называют рауберовым слоем (рис. 39, 4). В ходе дальнейшего развития зародышевый узелок превращается в зародыше-
158 |
Болотов А. В. Биология размножения и развития |
вый щиток (рис. 39, 7). Начинаются процессы гаструляции. В зародышевом щитке обособляется энтодерма, в результате чего он становится двухслойным, состоящим из эпи- и гипобласта. В зародышевом щитке, как и у птиц, будут происходить процессы формирования первичной полоски и гензеновского узелка (рис. 42), т. е. тех же структур, с которыми связано образование системы осевых органов зародыша.
На стадии гаструляции зародыш имплантируется в стенку матки. Это – сложный в морфологическом и физиологическом отношениях процесс. Зародыш внедряется в слизистую оболочку матки целиком или только отростками трофобласта. Выросты трофобласта называются первичными ворсинками. Впоследствии развиваются вторичные ворсинки с соединительной тканью и сосудами. Так формируется хорион – ворсинчатая оболочка, гомологом которой является серозная оболочка зародышей рептилий и птиц.
Рис. 39. Разрез ранней (А) и поздней (Б) бластоциты и дифференцирующего эмбриобласта (В) (по К. Уоддингтону, 1957).
1 – трофобласт, 2 – эмбриобласт, 3 – зародышевый узелок, 4 – рауберов слой трофобласта, 5 – эктобласт (эпибласт), 6 – энтодерма (гипобласт), 7 – зародышевый щиток
X. ПРОВИЗОРНЫЕ (ВНЕЗАРОДЫШЕВЫЕ) ОРГАНЫ
Провизорные, или временные, органы, развивающиеся в процессе эмбриогенеза вне тела зародыша, выполняют многообразные функции, обеспечивающие рост и развитие самого зародыша. В связи с тем, что некоторые из этих органов окружают зародыш, распространено и другое название – зародышевые оболочки. К ним относят: желточный мешок, амнион. серозную оболочки, аллантоис, хорион, плаценту. В эволюции они появились не одновременно.
Желточный мешок
В ряду хордовых животных внезародышевые органы впервые появляются у рыб в виде желточного мешка, депонирующего желток, используемый зародышем в процессе развития. Его формирование начинается на стадии ранней гаструлы, когда во внутреннем листке можно выделить зародышевую (кишечную) энтодерму и расположенную по периферии диска внезародышевую желточную энтодерму. Своим свободным краем желточная энтодерма образует край обрастания, который начинает надвигаться на желток. После возникновения хордомезодермального зачатка между экто- и энтодермой прорастают париетальный и висцеральный листки мезодермы. Желток обрастает всеми четырьмя листками. Зародыш приподнимается над диском и отделяется от желтка туловищной складкой. При образовании туловищной складки зародышевая энтодерма, до того распластанная на желтке, сворачивается в кишечную трубку. Зародыш связан с желточным мешком полым канатиком – желточным стебельком. Желточный мешок рыб выполняет трофическую функцию. Другая функция мешка – кроветворная – заключается в образовании клеток крови в мезодерме стенки мешка.
Свыходом животных на сушу (у пресмыкающихся и птиц)
всвязи с развитием зародыша под скорлупой появляются новые зародышевые органы: амнион, серозная оболочка и аллантоис.
160 |
Болотов А. В. Биология размножения и развития |
Как и у рыб, у пресмыкающихся и птиц возникают туловищные складки, отделяющие зародыш от желточного мешка. Желточный мешок у них также выполняет трофическую и кроветворную функции.
Амнион
Позднее в эмбриогенезе пресмыкающихся и птиц за счет эктодермы и париетального листка мезодермы формируются амниотические складки, растущие в направлении дорсальной поверхности зародыша. По мере роста головного конца зародыша амниотические складки как бы надвигаются спереди на зародыш, причем он одновременно вдавливается в желток (рис. 40, А, Б). Нарастающие на зародыш складки смыкаются и оба листка – эктодерма и прилегающий к нему париетальный листок мезодермы срастаются с одноименными листками противоположной стороны. Из двух листков складок при этом образуются две оболочки – амниотическая, или водная, обращенная к зародышу, и серозная, наружная (рис. 40, В).
Рис. 40. Схема развития внезародышевых органов у млекопитающих.
А – сближение амниотических складок и начало образования аллантоиса; Б, В – сформированные внезародышевые органы; 1 – тело зародыша; 2 – туловищная складка; 3 – амниотические складки; 4 – полость амниона; 5 – желточный мешок; 6 – аллантоис; 7 – амниотическая оболочка; 8 – кишка; 9 – хорион
Амниотическая оболочка на ранних стадиях отделена от тела зародыша узкой щелью, которая позднее превращается в за-