

4 КУРС (ЭМБРИОЛОГИЯ) / Несортированное по эмбриологии / ТАБЛИЦА
.pdfVI. Оплодотворение |
131 |
мартин) – и бычок. Это происходило из-за того, что гормоны мужского эмбриона начинали синтезироваться раньше, чем гормоны женского эмбриона.
Унекоторых видов происходит переопределение пола. Так,
вТихом океане обитают рыбки Labroides dimidiatus, живущие стайками из самок и одного самца. Каждый член кроме самца находится в состоянии стресса, источником которого является сам самец. При этом можно выделить разные уровни самок. Гибель самца сбрасывает напряжение с главной самки, и она становится самцом. Описанное явление зависит от гормонов выделяемых клетками надпочечников.
Уморского червя Bonella viridis пол зависит от того, будет зигота развиваться в непосредственном контакте с материнским организмом или самостоятельно.
Искусственное изменение условий среды
Проводилось с рыбами, амфибиями, птицами. Если кормить мальков самцов аквариумных рыб до 8 месяцев кормом с добавлением женских половых гормонов (эстрогенов), приводило к превращению их в самок.
Новые данные о детерминации пола у птиц
Недавние исследования на гинандроморфных (у которых одна половина мужская, а другая – женская; это крайне редкое природное отклонение) цыплятах дали неожиданный результат (рис. 26). Оказалось, что эти животные – истинные химеры, то есть представляют собой «коктейль» из клеток, несущих хромосомы ZZ (определяющие мужской фенотип у птиц), и ZW (определяющие женский фенотип).
Эти данные посеяли сомнения в том, что для птиц верна классическая модель детерминации пола, когда фенотип большей частью определяется гормональной секрецией половых желез, а те, в свою очередь, начинают развиваться в ответ на активацию некоего «полового» гена (например, у млекопитающих такой ген называется Sry и определяет развитие яичек). Если же организм цыпленка, несмотря на общий гормональный фон, разделяется на две разнополые половины, это говорит о том, что соматические клетки птиц определяют свой пол автономно, а не генерализованно, как считалось раньше.

132 |
|
Болотов А. В. Биология размножения и развития |
|||
|
|
|
|
Чтобы проверить это предпо- |
|
|
|
|
|
ложение, были проведены экспе- |
|
|
|
|
|
рименты на искусственных химе- |
|
|
|
|
|
рах. Для этого мезодермой двух- |
|
|
|
|
|
дневных GFP-эмбрионов, нахо- |
|
|
|
|
|
дящихся на 12-й стадии развития, |
|
|
|
|
|
заменяли аналогичный участок в |
|
|
|
|
|
левой части не GFP-эмбрионов |
|
|
|
|
|
такого же возраста (поскольку у |
|
|
|
|
|
птиц полностью развивается толь- |
|
|
|
|
|
ко левый яичник). На 35-й стадии |
|
|
|
|
|
развития (9 дней инкубации) эм- |
|
|
|
|
|
брионы исследовали и обнаружи- |
|
|
|
|
|
ли (с помощью иммуногистохи- |
|
|
|
|
|
мии), что в разнополых химерах |
|
|
|
|
|
донорские клетки хранят верность |
|
|
|
|
|
своему первоначальному полу: то |
|
|
|
|
|
есть мужские клетки, будучи по- |
|
|
|
|
|
мещенными в женское общество, |
|
|
|
|
|
не начинают вырабатывать арома- |
|
|
|
|
|
тазу, а женские, оказавшиеся в |
|
|
|
|
|
мужской гонаде, – наоборот, не |
|
|
|
|
|
вырабатывают |
анти-мюллеров |
|
|
|
|
гормон. Более того, одна из полу- |
|
|
|
|
|
ченных химер образовала яични- |
|
|
|
||||
ко-яичко – то есть передняя его часть состояла из женских клеток и экспрессировала ароматазу, а задняя– из мужских клеток и вырабатывала анти-мюллеров гормон, что стало дополнительным свидетельством автономнойполовойдетерминации клетками птиц.
VII. ДРОБЛЕНИЕ
Оплодотворение играет чрезвычайно важную роль в развитии, но это лишь его первая ступень. Зигота ее генетическим материалом и определенным распределением цитоплазмы приступает к созданию многоклеточного организма. У всех известных животных это наблюдается в процессе дробления – серии митотических делений, в результате которых объем цитоплазмы яйца разделяется на клетки меньшего размера, которые называются бластомерами.
Убольшинства видов в период дробления объем зародыша не увеличивается и огромная масса цитоплазмы зиготы разделяется на всё более мелкие клетки. Это деление без роста объема сопровождается выпадением интерфазного периода роста между делениями, тогда как деление ядер следует друг за другом с большой скоростью, которая никогда не наблюдается в дальнейшем. Одним из факторов, влияющих на скорость делений является соотношение ядра и цитоплазмы, существенно отличающееся от такового у соматических клеток. В процессе делений дробления скорость их постепенно уменьшается. Таким образом дробление начинается сразу после оплодотворения и заканчивается, когда у зародыша достигается новое равновесие между ядром и цитоплазмой, соответствующее соматическим клеткам.
На ранних стадиях деления дробления фаза S занимает очень маленькое время (≈10 мин), а в дальнейшем оно растет как
впроцессе дробления, так и в общем эмбриогенезе. Например, в период гаструляции фаза S ≈ 1 ч, а во время нейруляции – 5 ч.
Унекоторые животные после завершения дробления наступает пауза, которая заканчивается в определенные сроки. У летучих мышей осеменение происходит поздней осенью, тогда как развитие яйца начинается весной. Пауза типична и для ракообразных.
Скорость дробления изменяется под влиянием температуры. Существует следующая закономерность: при изменении температуры на 10 ºС деление дробления ускоряется (замедляется) в 2–3 раза. Ранний период развития характеризуется синхронностью деления. Возможно, что деление дробления является еди-
134 |
Болотов А. В. Биология размножения и развития |
ницей для регуляции продолжительности различных периодов развития в дальнейшем. Поздний период дробления называется бластуляцией. Он характеризуется: 1) удлинением фазы S; 2) ускорением синтетических процессов. Геном зародыша начинает функционировать не сразу, а после прохождении определенного числа делений дробления и нормализации ядерно-плазменных отношений.
По мере дробления включаются факторы взаимодействия бластомеров, факторы интеграции зародыша, а это ведет к тому, что темпы деления клеток в разных частях зародыша становятся неодинаковыми. Угнетение одних частей зародыша на ранних стадиях онтогенеза.
Если темпы дробления сохранились бы прежними и дробление было бы синхронным, то через 20 ч развития яйца, например, лягушки (начало гаструляции) зародыш бы состоял из 1 млн клеток, а через 2 суток – 25·1013 клеток. К моменту ранней хвостовой почки (к концу седьмых суток) количество клеток было более 1028, а размер зародыша оказался бы 5,2·1012 м3. И это было бы при диаметре клеток ≈ 10 мкм.
В 1962 г. Уильям С. Буллоу в межклеточной среде обнаружил вещества, концентрация которых подавляет деление клеток. Они были названы кейлоны (англ. кеу – ключ, long – единственный), которые тормозят в окружающих клетках реакции митоза, скорость синтеза нуклеиновых кислот и белков, и при этом не обладают видовой специфичностью. Концентрация этих веществ зависит от числа клеток в данном объёме, а количество кейлона, вырабатываемое одной клеткой является величиной постоянной.
Итак, дробление – строго координированный процесс, находящийся под генетическим контролем. Образование многоклеточности – первая и основная функция дробления. Другая функция – в увеличении ядерно-плазменного отношения.
Видовые особенности этого процесса определяются двумя основными параметрами:
1)количеством и распределением желтка в цитоплазме;
2)присутствием в цитоплазме факторов, влияющих на ориентацию митотического веретена и время его образования.
Если желток в яйце распределен неравномерно, то в области, содержащей меньше желтка (анимальный полюс), клеточные
VII. Дробление |
135 |
деления идут с большей скоростью, чем в противоположной богатой желтком области (вегетативный полюс). Ядро зиготы в этом случае сдвигается к анимальному полюсу. То есть можно сказать следующее – желток подавляет дробление.
Правила зависимости между расположением желтка и положением ядер и веретен были сформулированы немецким ученым О. Гертвигом на основе правил, предложенных ранее ботаником Ю. Саксом для растительных меристем. Эти правила состоят в следующем: 1) клеточное ядро стремится расположиться в центре свободной от желтка цитоплазмы; 2) веретено клеточного деления стремится расположиться по направлению наибольшего протяжения свободной от желтка цитоплазмы.
Перетяжки, разделяющие дробящуюся зиготу на всё более мелкие клетки (бластомеры), называют бороздами дробления. Борозды дробления делят на следующие виды:
1)меридианная (меридиональная) – проходит через ани- мально-вегетативную ось зиготы;
2)экваториальная, которая может быть смещена к анимальному полюсу из-за большого количества желтка в вегетативном полюсе, проходит по широте зиготы (поперек) или по экватору зиготы;
3)тангенциальная – проходит параллельно поверхности зиготы, так образуется несколько слоев клеток зародыша.
У зигот, содержащих относительно мало желтка (изолецитальные, мезолецитальные яйца), в процессе деления борозда дробления проходит через всё яйцо, то говорят, что дробление полное, или голобластическое.
Зиготы, содержащие много желтка (полилицетальные яйца)
ипо расположению желтка находящегося в вегетативной области (телолецитальные яйца), в процессе деления борозды дробления не проникают в богатую желтком область цитоплазмы, т. е. не проходят через все яйцо, в этом случае дробление неполное или меробластическое.
Однако желток – только один из факторов, оказывающий влияние на характер дробления зиготы. Существуют также унаследованные особенности клеточных делений, которые накладываются на влияние желтка. Это можно увидеть на примере изолецитальных яиц, содержащих очень мало желтка.
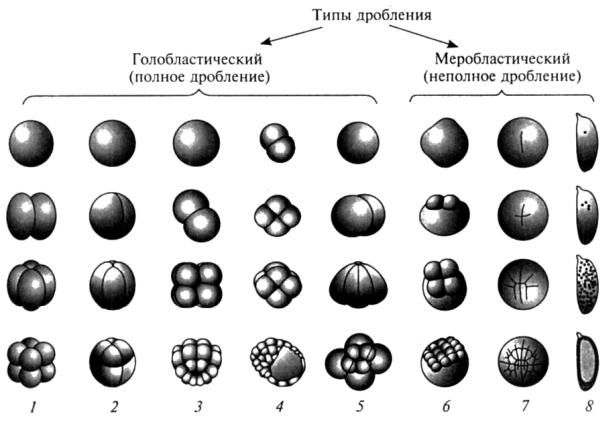
136 |
Болотов А. В. Биология размножения и развития |
При отсутствии большого количества желтка наблюдается четыре типа дробления (рис. 27, 1–5):
Рис. 27. Классификациятиповдробления(поГоличенковуВ. А., 2004): 1 – полное равномерное дробление (иглокожие, бесчерепные); 2 – полное неравномерное дробление (амфибии, осетровые рыбы); 3 – полное билатеральное дробление (асцидии); 4 –полное равномерное ротационное дробление (плацентарные млекопитающие); 5 – полное спиральное дробление (большинство моллюсков; кольчатые, плоские и круглые черви); 6, 7 – неполное дискоидальное дробление (6 – костистые рыбы; 7 – репти-
лии, птицы); 8 – неполное поверхностное дробление (насекомые)
1. Радиальное голобластическое дробление, которое может быть 2 видов:
а) полное равномерное дробление (бесчерепные (ланцетник) и иглокожие). Олиголецитальные яйцеклетки (рис. 27, 1);
б) полное неравномерное дробление (амфибии и осетровые). Мезолецитальные яйцеклетки) (рис. 27, 2).
2.Спиральное голобластическое дробление (большинство моллюсков, кольчатые, круглые и плоские черви) (рис. 27, 5).
3.Билатеральное голобластическое дробление (асцидии) (рис. 27, 3).
VII. Дробление |
137 |
4. Асинхронное, ротационное голобластическое дробление (плацентарные млекопитающие) (рис. 27, 4).
При наличии большого количества желтка наблюдается два типа дробления (рис. 27, 6–8):
1)дискоидальное меробластическое дробление (рептилии, птицы, костистые рыбы) (рис. 27, 6–7);
2)поверхностное меробластическое дробление (насекомые) (рис. 27, 8).
1. Радиальное голобластическое дробление
Полное равномерное дробление
Указанный тип дробления встречается у иглокожих и бесчерепных (ланцетник). Их яйцеклетка – алецитальная или первично олиго- и изолецитальная, т. е. не содержит вовсе или содержит очень мало желтка (распределенного равномерно).
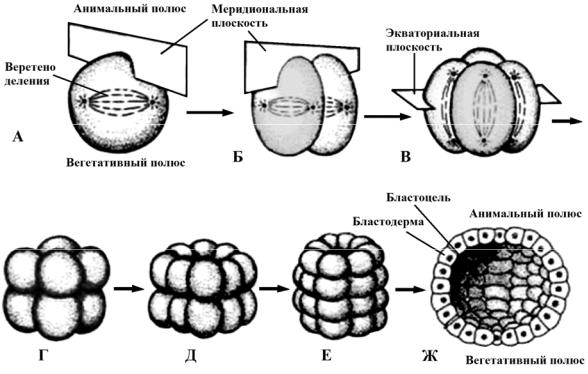
Поэтому дробление полное (дробятся все части зиготы), равномерное (образуются клетки одинаковой величины) и синхронное (деления разных клеток зародыша происходят одновременно) (рис. 28, А–Е).
Борозды дробления ориентированы параллельно или перпендикулярно анимально-вегетативной оси. После соединения пронуклеусов ось первого митотического веретена ориентируется перпендикулярно анимально-вегетативной оси яйца. Первая борозда дробления проходит через анимальный и вегетативный полюсы яйца, т. е. через анимально-вегетативную ось и ориентированы вдоль неё. Такое деление называют еще меридиональным, поскольку борозда деления проходит через два полюса подобно меридиану на глобусе (рис. 28, А). Это приводит к образованию двух одинаковых дочерних клеток. Плоскость второго деления тоже проходит через оба полюса, но перпендикулярно плоскости первого деления (рис. 28, Б). Таким образом, два первых деления дробления меридиональны, но взаимно перпендикулярны. Третье деление экваториальное, т. е. митотические веретёна в каждом бластомере располагаются параллельно ани- мально-вегетативной оси (рис. 28, В).
Все образующиеся бластомеры одинаковы по размерам. Четвертое деление снова меридиональное и приводит к образованию двух ярусов по 8 клеток в каждом. Пятое – экваториальное.
138 |
Болотов А. В. Биология размножения и развития |
Рис. 28. Радиальное голобластическое равномерное дробление (А, Б, В, Г, Д, Е) (ланцетник) и образование бластулы (Ж)
Дальше наблюдается чередование меридиональных и экваториальных делений. В результате образуется зародыш, у которого бластомеры располагаются горизонтальными рядами. На обоих полюсах зародыша бластомеры сближаются и возникает шар с полостью внутри. Такой шар называется бластулой, полость бластоцель. Если разделить зародыш по меридиану или по экватору, то образуются две зеркальные половинки. Такой тип симметрии присущ шару и носит название радиальной симметрией. Образовавшаяся бластула называется целобластула: ее стенка – бластодерма – составляет один слой одинаковых по размеру клеток, а полость – бластоцель – расположена в центре (рис. 28, Ж). По существу, это однослойный сферический пузырек, стенка которого везде одинакова по толщине.
Полное неравномерное дробление
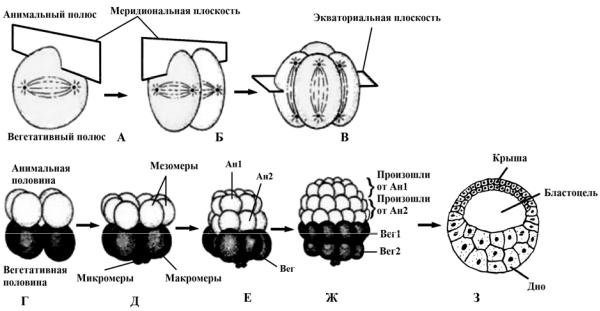
Для амфибий и осетровых рыб также характерно радиальное голобластическое дробление, но с существенными модификациями (рис. 29). Яйцеклетка – умеренно телолецитальная: желтка много, и он в большей мере сосредоточен у вегетативного полюса яйцеклетки. Это придает дроблению некоторые особенности: дробление полное (дробятся все части зиготы), но при
VII. Дробление |
139 |
этом неравномерное (на вегетативном полюсе зародыша клетки крупнее), и асинхронное (клетки вегетативного полюса делятся медленней).
Рис. 29. Полное неравномерное дробление (А, Б, В, Г, Д, Е, Ж) и образование амфибластулы (З)
Первое и второе деление проходят так же, как и у иглокожих и бесчерепных (рис. 29, А, Б). Третье деление тоже экваториальное и отделяет один полюс от другого, плоскость этого деления смещена в анимальное полушарие, поэтому дробление получило характеристику неравномерное, ведущее к образованию микромеров в анимальном и макромеров в вегетативном полушариях (рис. 29, В, Г). На анимальном полюсе четыре более мелких бластомера – микромеры, на вегетативном – четыре более крупных – макромеры (рис. 29, Д). Затем деление опять идет в меридианных плоскостях, а потом опять в широтных и т. д. У амфибий первые четыре деления проходят синхронно, последующие 7 делений волнами, начинающимися на анимальном полюсе и заканчивющиеся на вегетативном (рис. 29, Ж).
В результате подобного дробления образуется амфибластула (рис. 29, З). Ее особенности следующие:
1)анимальный полюс, называемый крышей, содержит мелкие клетки;
2)вегетативный полюс, или дно, состоит из крупных клеток;
140 |
Болотов А. В. Биология размножения и развития |
3)область же между крышей и дном – краевая зона, клетки здесь – промежуточного размера;
4)бластодерма во всех перечисленных областях является
многослойной;
5)бластоцель смещена к крыше.
2. Спиральное голобластическое дробление
Спиральное дробление характерно для нескольких групп червей (кольчатых, круглых и плоских) и почти всех моллюсков, за исключением головоногих. Оно во многих отношениях отличается от радиального дробления.
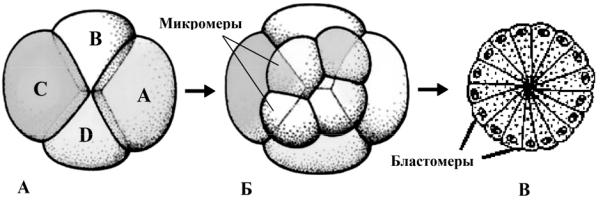
Во-первых, яйца не делятся параллельно или перпендикулярно анимально-вегетативной оси зиготы. Плоскости делений дробления ориентированы наклонно (под некоторым углом), что и приводит к «спиральному расположению дочерних бластомеров (рис. 30).
Во-вторых, число контактов между клетками больше, чем при радиальном дроблении. Бластомеры принимают расположение, наиболее стабильное в термодинамическом отношении, и минимальным объёмом, подобно расположению контактирующих мыльных пузырей.
Рис. 30. Спиральное дробление (А, Б) и образование стерробластулы (В)
В-третьих, зародыш с этим типом дробления проходит меньшее число делений до начала гаструляции. Это позволило проследить судьбу каждой отдельной клетки бластулы. Когда сопоставили судьбу отдельных клеток зародышей кольчатых