

Цель занятия:
1. Научиться объяснять микроэлектродный метод измерения биопотенциалов.
2. Научиться объяснять в рамках электродиффузионной теории природу потенциала покоя и уметь рассчитать его величину по заданным концентрациям ионов.
3. Научиться объяснять процесс генерации потенциала действия. Уметь начертить его график.
4. Научиться объяснять и изображать графически ионные потоки через мембрану в процессе возбуждения клетки.
ПЛАН ИЗУЧЕНИЯ ТЕМЫ:
ИСХОДНЫе ЗНАНИя:
1. Знать природу электрического потенциала и разности потенциалов.
2. Знать их единицы измерения.
ПЛАН ИЗУЧЕНИЯ ТЕМЫ:
1. Биоэлектрические потенциалы. Мембранные потенциалы и их ионная природа.
2. Микроэлектродный метод внутриклеточного измерения потенциалов.
3. Потенциал покоя в клетках.
4. Потенциал действия. Свойства потенциала действия.
5. Механизм генерации и распространения потенциала действия.
6. Опыты с фиксацией напряжения.
7. Распространение потенциала действия вдоль возбудимого волокна.
Литература:
1. Лекции.
2. В.Ф. Антонов и др. Биофизика, М. Владос, 2000. гл. 5, с.123 - 127.
3. В.Ф. Антонов и др. Практикум по биофизике, М. Владос, гл. 2,. с. 80-86.
2. Рубин А.Б. Биофизика, М. Высшая школа, 1987. т. 2.
ТЕОРЕТИЧЕСКие предпосылки работы
В результате жизнедеятельности в клетках и тканях могут возникать разности электрических потенциалов: окислительно-восстановительные и мембранные. Биопотенциалы, регистрируемые в организме, - это в основном мембранные потенциалы.
Если концентрация какого-либо иона внутри клетки отличается от концентрации этого иона снаружи и мембрана проницаема для этого иона, возникает поток заряженных частиц. Математически поток ионов описывается уравнением Нернста-Планка:
Iм = –uRT (dc/dx) – uczF (dφ/dx), (1)
где u — подвижность иона, R — газовая постоянная, Т — абсолютная температура, с —концентрация ионов, z — заряд иона в элементарных единицах заряда, F — постоянная Фарадея, dc/dx и dφ/dx — градиенты концентрации и потенциала. В состоянии покоя проницаемость мембраны для ионов К+ значительно больше, чем для Na+, и больше, чем для Cl-. В начальный момент φвн = φнар поэтому dφ/dx = 0. Таким образом, в начальный момент поток ионов К+ из клетки наружу Iнар определяется только диффузией: Iнар = -uRT (dc/dx).
По мере выхода
ионов K+
из клетки второе слагаемое уравнения
(1) увеличивается до тех пор, пока: │uRT
(dc/dx)
│ = │uczF
(dφ/dx)
│. Тогда Iм,
т.е. в состоянии покоя (когда клетка не
возбуждена) ток через мембрану не идет.
Возникающая при этом разность потенциалов
на мембране является потенциалом покоя
![]() ,
величину которого можно оценить по
формуле Нернста:
,
величину которого можно оценить по
формуле Нернста:
![]()
Потенциалом действия (ПД) называется электрический импульс, обусловленный изменением ионной проницаемости мембраны и связанный с распространением по нервам и мышцам волны возбуждения. Все раздражители, действующие на клетку, вызывают в первую очередь снижение ПП; когда оно достигает критического значения (порога), возникает активный распространяющийся ответ – ПД. На рисунке 1 представлен ПД гигантского аксона кальмара.
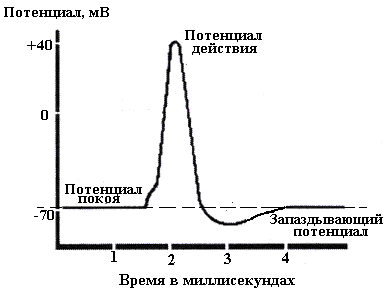
Рис.1.
Во время восходящей фазы ПД аксона внутренняя сторона мембраны, заряженная в покое электроотрицательно, приобретает в это время положительный потенциал. Достигнув вершины, ПД начинает падать (нисходящая фаза ПД), и потенциал на мембране возвращается к уровню, близкому к исходному, - ПП. Для ПД аксона характкрен так называемый запаздывающий потенциал, когда в течение некоторого времени на мембране существует даже меньший потенциал, чем потенциал покоя. Согласно мембранной теории, деполяризация мембраны, вызванная действием раздражителя, приводит к усилению потока Na+ внутрь клетки, что уменьшает отрицательный потенциал внутренней стороны мембраны - усиливает её деполяризацию. Это, в свою очередь, вызывает дальнейшее повышение проницаемости для Na+ и новое усиление деполяризации и т.д. Повышение проницаемости для Na+ очень кратковременно и сменяется её падением, а следовательно, уменьшением потока Na+ внутрь клетки. Проницаемость для К+, в отличие от проницаемости для Na+, продолжает увеличиваться, что приводит к усилению потока К+ из клетки. В результате этих изменений ПД начинает падать, что ведёт к восстановлению ПП. Таков механизм генерации ПД в большинстве возбудимых тканей.
Своеобразен механизм генерации ПД в мышечных волокнах сердца, для которых характерно длительное плато на нисходящей фазе ПД. Неравенство концентраций ионов К+ и Na+ (или Ca+) внутри и снаружи клетки (волокна) поддерживается специальным механизмом (т. н. "натриевым насосом"), выталкивающим ионы Na+ из клетки и нагнетающим ионы К+ в протоплазму, требующим затраты энергии, которая черпается клеткой в процессах обмена веществ.
Амплитуда ПД большинства нервных и мышечных волокон примерно одинакова: 110-120 мВ. Длительность ПД варьирует в широких пределах: у теплокровных животных длительность ПД нервных волокон, наиболее быстро проводящих возбуждение, - 0,3-0,4 мс, у волокон же мышц сердца - 50-600 мсек. Характерной особенностью ПД, отличающей его от других форм ответа клетки на раздражение, является то, что он подчиняется правилу "всё или ничего", т. е. возникает только при достижении раздражителем некоторого порогового значения, и дальнейшее увеличение интенсивности раздражителя уже не сказывается ни на амплитуде, ни на продолжительности ПД.
Потенциал действия - один из важнейших компонентов процесса возбуждения. В нервных волокнах он обеспечивает проведение возбуждения от чувствительных окончаний (рецепторов) к телу нервной клетки и от неё - к синаптическим окончаниям, расположенным на различных нервных, мышечных или железистых клетках. Поступая в эффекторные окончания, ПД вызывает выделение (секрецию) определённой порции специфических химических веществ, т. н. медиаторов, оказывающих возбуждающее или тормозящее влияние на соответствующие клетки. В мышечных волокнах распространяющийся ПД вызывает цепь физико-химических реакций, лежащих в основе процесса сокращения мышц.
Вопросы для самоконтроля:
1. Что называется трансмембранной разностью потенциалов?
2. Каким способом можно измерить трансмембранный потенциал в эксперименте?
3. Как формируется потенциал покоя клетки? Каковы соотношения основных ионов на внешней и внутренней стороне мембраны?
4. Понятие об ионных каналах мембраны и их характеристика.
4. Уравнение Нернста для потенциала покоя.
5. Уравнение Гольдмана-Ходжкина -Катца.
5. Потенциал действия аксона. Основные свойства потенциала действия аксона.
6. Опыты с фиксацией напряжения.
7. Распространение потенциала действия по нервному волокну аксона кальмара и человека.
8. Механизм генерации потенциала действия кардиомиоцита.
Приборы и принадлежности:
Персональный компьютер.
Схема работы:
Последовательность действий |
Способ выполнения задания |
1. Изучение микроэлектродного метода регистрации трансмембранного потенциала Δφм. |
1. Выполните задание №1. 2. Ответьте на вопросы. Ответы занесите в тетрадь. |
2. Изучение процессов диффузии через каналы мембраны и возникающую при этом трансмембранную разность потенциалов |
1. 1. Выполните задание №2. 2. Ответьте на вопросы. Ответы занесите в тетрадь.
|
3. Изучение выравнивания электрохимического потенциала для ионов К+. |
1. Выполните задание №3. 2. Ответьте на вопросы. Ответы занесите в тетрадь. |
4. Изучение ионного состава системы клетка-внеклеточная жидкость при развитии потенциала действия. |
1. Выполните задание №4. 2. Ответьте на вопросы. Ответы занесите в тетрадь. |
5. Изучение кинетики токов через биологическую мембрану в процессе возбуждения. |
1. Выполните задание №5. 2. Ответьте на вопросы. Ответы занесите в тетрадь. |