

Поширення потенціалу дії. Кабельна теорія
Нервові та м'язові волокна є циліндричними провідниками, їхній внутрішній вміст, що має відносно низький питомий опір (за даними різних авторів, він може становити від 30 до 200 Ом • см), мембраною ізольовано від зовнішнього високо провідного середовища. Відповідно до орієнтовних розрахунків Ходжкіна, питомий опір комплексу структур, що утворюють мембрану, досягає 2 • 109 Ом • см. У стані спокою нервові й м'язові волокна проводять електричний струм, як коаксіальний кабель.
Електрична ємність плазматичної мембрани різноманітних клітин дорівнює приблизно 1 мкФ/см2. Електрична ємність чистого бімолекулярного ліпідного прошарку становить 0,8 мкФ/см2.
Знаючи діелектричну проникність тієї частини ліпідного бішару, що складається з вуглеводневих ланцюгів, можна розрахувати товщину ізолюючої частини d, використавши формулу плоского конденсатора:
![]()
де С- електрична ємність; ε - діелектрична проникність ізолюючої частини бішару; εо - електрична стала; S - площа. Якщо С = 0,8 мкФ/см2, а ε = 2, то d = 2,2 нм.
Електрична ємність біологічних мембран є важливим параметром, який слід ураховувати при кількісній оцінці електричних сигналів, що реєструються на мембрані. Знаючи ємність мембрани, можна розрахувати мінімальну кількість зарядів (іонів), необхідних для виникнення певної зміни мембранного потенціалу. Вимірювання електричної ємності плазматичної мембрани дозволяє орієнтовно оцінити площу поверхні клітини.
Волокно можна уявити як електричний ланцюг з розподіленими параметрами. Для зручності розгляду й кількісного опису такого ланцюга припускають, що коли волокно занурене у великий об'єм рідини, то провідність зовнішнього середовища є достатньо великою і в еквівалентному електричному ланцюзі не враховують його опір. На рис. 9.3, а наведено схему кабельної структури. Опір мембрани й цитоплазми, а також електричну ємність мембрани зображено на схемі у вигляді окремих повторюваних елементів. Потенціал позаклітинного середовища розглядають як нульовий. Падіння напруги Е на опорі мембрани у разі проходженні струму через кабельну структуру називають електротонічним потенціалом.
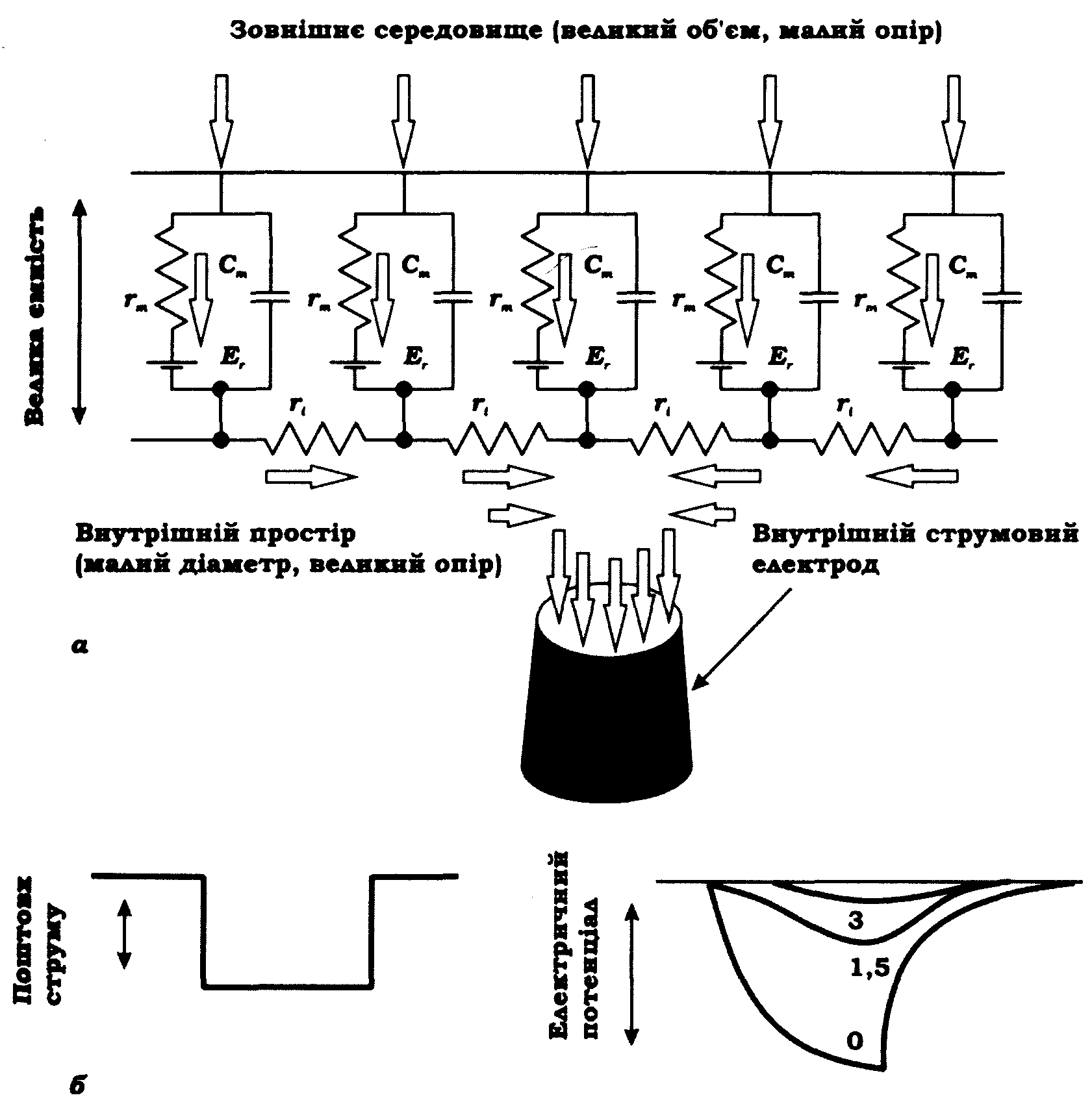
Рис. 9.3. Кабельна структура (К. Катц, 1966):
а - електрична схема, що ілюструє розподіл опору та ємності мембрани, а також опору протоплазми (стрілки вказують напрямок струму);
б - прямокутний поштовх струму й зумовлені ним зміни мембранного потенціалу на зазначених відстанях від електрода (відстані подано у константах довжини λ)
Вираз для електротонічного потенціалу Е на відстані х має наступний вигляд:
![]()
де E0 – потенціал при х = 0; rm - опір мембрани кабельної структури, розрахований на одиницю довжини; ri – опір аксоплазми, розрахований на одиницю довжини осьового циліндра.
Цей вираз доводить, що електротонічний потенціал Е експоненціальне затухає зі збільшенням відстані х. Відстань х, на якій електротонічний потенціал затухає в е разів, називають константою довжини й позначають як λ = (rm / ri )1/2.
Розглядаючи залежності λ, від параметрів кабельної структури, rm виражають через Rт - опір мембрани, розрахований на одиницю площі, а ri через Ri, - питомий опір цитоплазми:

де D - діаметр волокна. Тоді константа довжини становить:
![]()
тобто вона є пропорційною кореню квадратному з діаметра волокна. У табл. 9.1 наведено кабельні параметри нервових волокон.
Таблиця 9.1. Кабельні параметри нервових волокон (за Т= 293 К)

Поширення потенціалу дії відбувається завдяки виникненню локальних струмів між збудженою та незбудженою ділянками нервового або м'язового волокна.
Початок потенціалу дії являє собою електротонічний процес (деполяризація мембрани до підпорогового рівня). На рис. 9.4 наведено теоретично розраховану схему Д. Нобла, яка ілюструє зміни мембранного потенціалу й локальні струми при поширенні потенціалу дії в кабельній структурі.
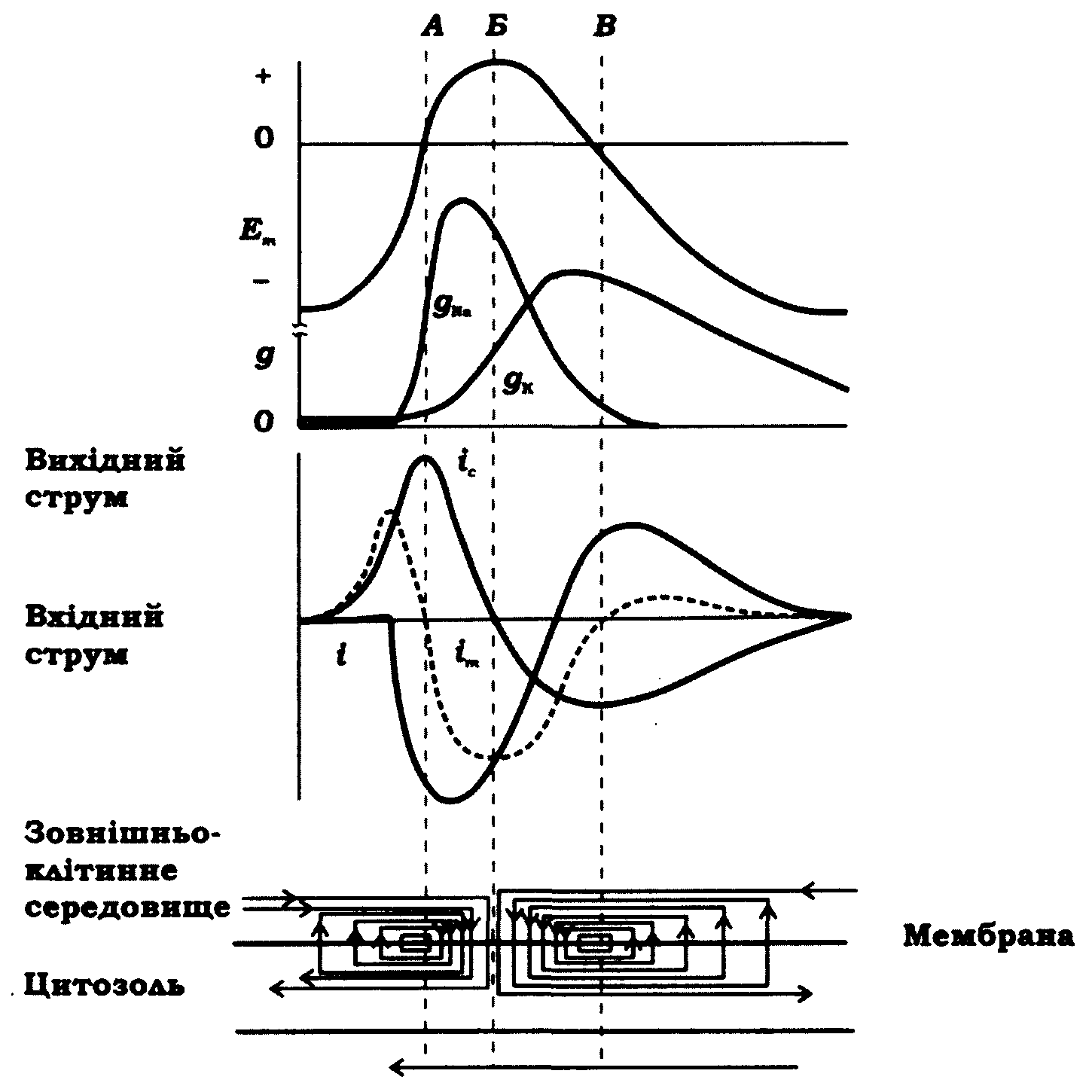
Рис. 9.4. Зміни мембранного потенціалу Еm, натрієвої та калієвої провідності мембрани вздовж кабельної структури під час поширення потенціалу дії (Д. Нобл, 1966):
іі - струм через іонні канали; іс - ємнісний струм; іт - сумарний струм (іт = іі + іс).
На нижній діаграмі зображено локальні струми в ділянці збудження. Потенціал дії поширюється ліворуч, як показано стрілкою. Вертикальні штрихові лінії дають можливість зіставляти події в місцях нульового рівня мембранного потенціалу та його максимуму, а також у місці зміни напрямку струму іт у спадаючій фазі потенціалу дії
Тут же подано криві, що характеризують трансмембранні струми (струм, що входить у волокно - негативний, а виходить з нього - позитивний), зміни мембранного потенціалу вздовж: волокна й провідності мембрани для іонів натрію та калію у визначені моменти потенціалу дії. Довжина ділянки волокна, де існує потенціал дії, залежить від швидкості поширення. Ділянка між лініями А і Б є повністю збудженою.
Вхідний іонний струм у волокно переносять іони натрію. Це супроводжується зміною мембранного потенціалу в місці збудження й зумовлює поздовжні струми, необхідні для електротонічної деполяризації мембрани незбуджених раніше ділянок до порогу виникнення потенціалу дії. Електротонічна деполяризація на початку потенціалу дії досягає порогового рівня, що приводить до збільшення gNa й виникнення збудження.
Наприкінці потенціалу дії праворуч від лінії В спостерігається також вихідний мембранний струм, що може викликати деполяризацію мембрани. Тут значна частина натрієвих каналів є інактивованою. Крім того, деполяризація шунтується високою калієвою провідністю мембрани. Ці обставини забезпечують рефрактерний стан.
Що більшим є внутрішній опір кабельної структури, її ємність й опір зовнішнього середовища, то меншою є швидкість проведення. Внутрішній опір змінюється в оберненій залежності від квадрата діаметра волокна. Швидкість поширення потенціалу дії є пропорційною кореню квадратному з діаметра волокна.
У мієлінізованих нервових волокнах потенціал дії (нервовий імпульс) виникає лише в перехопленнях Ранв'є. Між перехопленнями нервове волокно вкрите мієліном, утвореним шванівскими клітинами. Мієлін є ізолятором, що надає можливість потенціалу дії перестрибувати з одного перехвату до іншого. Таке проведення нервового імпульсу має назву стрибкоподібного (сальтоторного).