

Органические вещества

Минеральные
соли
Источники
микроэлементы
азота
Рис.
60.
Схема метаболизма бактерий
АНАБОЛИЗМ
КАТАБОЛИЗМ
Мономеры
(строительные белки)
Аминокислоты
Сахара
Нуклеотиды
Рис.
61.
Катаболизм и анаболизм клеток
Полимеры
( макромолекулы ) Белка
Полисахариды
Полииуклеотнды
©
фруктозо-1,6-бифосфатный
путь Пентаэофосфатный путь
§2-кето-3-пе»кси-6-фосфогл»оконатиый
путь Цикл трикарбоноаых кислот
Дыхательная цепь V (б^ Фосфорилироеание
на уровне субстрата
([.
Д /5ч Окислительное фосфорилироеание
I
HjOl в
дыхательной цепи
—'
(в) Синтез мономеров
Синтез
полимеров
для которых имеются соответствующие транспортные системы. Вещества могут проникать в клетку при пассивной диффузии. Таким путем в клетку в небольшом количестве проникают чужеродные вещества. Сахара, аминокислоты и другие метаболиты таким путем в клетку не проникают. При облегченной диффузии вещество транспортируется в клетку при условии, что его концентрация в клетке меньше, чем в окружающей среде. Этот процесс осуществляется субстрат-специфическими пермеазами и не требует затрат энергии. Активный транспорт требует затрат энергии в виде АТФ и вещество может накапливаться в клетке против градиента концентрации. При транслокации группы переносимая молекула в процессе транспорта видоизменяется, например фосфорилируется.
Существует принцип биохимического единства жизни на Земле, который заключается в универсальности ряда процессов и явлений. Так, универсален генетический код (ДНК, РНК), «строительные блоки». Например, белки состоят из аминокислот, нуклеиновые кислоты — из пуриновых и пиримидиновых оснований,
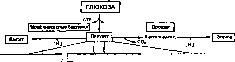
|глкжтл|
Н2(>
Пенто>офосфатный путь
Глюкозо-6-фосфот~|

(АТР)-^
Гексокинта
2-
Kemo-J-detoKL'M-6-
фосфоглюконатный
путь
Г
Глюкто-6-фосфаташ

Гекстобифосфаташ
н,о
Фруктозо-1,6-бнфосфат
^АТР
6-фосфофруктокинта
(ADP)
Фруктоюбифосфаашлкикигш
Триопофосфшп-томерта
Глниеральдегид-З-фосфат
/
лицеральдсгидфосфат- дегидрогенит
Фосфоглицераткина
ш
ГЙАОЙ^)
(NAD>\|

Баланс:
Глюкоза 2 Пируват + 2АТР + 2 NADH;
Рис. 62. Фруктозо-/,6-бифосфатный путь расщепления глюкозы (гликолиз)
полисахариды — из моиосахаров. Основной источник энергии (АТФ), пути расщепления Сахаров и дыхательная цепь едины. На фоне этого единства у микроорганизмов прослеживается много-
HjO
C.H3-CO-SC0A I СоА
CH2-COOH
НО-С-СООН
CHj-COOH Цитрат
СО-СООН
_
CHj-COOH
Оксалоацетат
®l/—-
(tfADF£)
/ (TTAED
н2о
2
с-соон
сн-соон
ццс-Аконитат
Н20
-сн-соон
I
сн2-соон
L
-Малат
V.®
®
®
н,о-
CHj-COOH
ОН,-СООН—СН-СООН I
CH.-COOH
I
2
©
нс-соон
II
ноос-сн
Фу
марат
СН2-СООН
сн2-соон
Сукиинат
V
сно-соон
Изоцитрат
(NAD(P)H
©
сн2-соон
сн-соон
I
CO-COOH
Оксалосукцинат
I 1
сн2
I
СоА
У\
СоА
(ADP)
сн2-соон
1
®
со^4
I
CO-COOH
2-Оксоглутарат
СоА
Рис. 63. Цикл трикарбоновых кислот
образие в процессах катаболизма и анаболизма, что выражается в различии источников энергии, источников углерода и восстановителя (водорода и электронов).

Наружная
сторона
простая
диффузия
Внутренняя
сторона
ФЕП
Фермент!
Пируват
Рис.
64.
Транспорт веществ в клетку
облегчённая
диффузия
активный
транспорт
транслокация
группы
По типам питания организмы могут быть разными. Главное — это источник энергии, донор водорода (электронов) и источник углерода для построения органического вещества:
источник энергии: а) фототрофы — источник энергии свет, б) хемотрофы — источник энергии химические вещества (органические или неорганические);
донор водорода (электронов): а) органотрофы используют органические вещества; б)литотрофы используют неорганические вещества — Н20, Н2, NH3, H2S, S, СО, Fe2+;
источник углерода: а) автотрофы используют С02, б) гете- ротрофы используют органические вещества.
При обозначении организма иногда используют только две, но чаще все три характеристики.
Примеры:
фотолитоавтотрофы — растения, цианобактерии, пурпурные и зеленые бактерии;
хемолитоавтотрофы — нитрификаторы, водородные бактерии, метаногенные бактерии;
хемоорганогетеротрофы — большинство бактерий, грибы, животные, человек.
Встречаются микроорганизмы, способные переходить от одного типа питания к другому, например, от фотоавтотрофии к хемогетеротрофии. Такие организмы называют миксотрофами. К ним можно отнести водоросли, например зеленую водоросль Chlorella vulgaris, которая может расти на свету за счет фотосинтеза, а при отсутствии света — за счет органических веществ.
Процессы катаболизма создают в клетке вещества с макроэр- гическими связями. Наиболее общее соединение такого типа — аденозинтрифосфат (АТФ), который образуется в результате реакций фосфорилирования. У микроорганизмов различают три типа фосфорилирования: 1) субстратное; 2) фотофосфорилиро- вание; 3) окислительное фосфорилирование. Появление этих типов реакций связано с ходом эволюции жизни на Земле. Если у высших организмов — растений и животных — преобладает один из типов энергетического метаболизма, то у более древних организмов прокариот имеются все указанные типы.
Наиболее примитивным способом получения энергии являются процессы субстратного фосфорилирования (фосфорилирования субстрата) — брожения, за счет которых существует несколько групп прокариот, широко представленных в почве. Брожение — это окислительно-восстановительные процессы превращения органических соединений, сопровождающиеся выходом энергии. Брожение протекает без участия молекулярного кислорода в анаэробных условиях, т.е. это окисление анаэробного типа. Электрон в окисляемых органических веществах переносится с одной части на другую часть органической молекулы, выступающей в роли его акцептора. Высвобождаемая на окислительном этапе процесса энергия запасается в молекулах АТФ. В среде накапливаются низкомолекулярные окисленные и восстановленные продукты — результат расщепления исходных молекул сбраживаемого субстрата. Брожению подвергаются углеводы, спирты, органические кислоты, аминокислоты, пурины и пиримидины. Продуктами брожения являются органические кислоты, спирты, ацетон, а также С02 и Н2. Обычно накапливается несколько продуктов, но по главному из них различают брожения: спиртовое, молочнокислое, маслянокислое и др. (рис. 65).
Следующим типом получения энергии вначале был также анаэробный процесс возникновения жизни за счет света— фото- фосфорилирование. В настоящий период существования жизни на нашей планете у микроорганизмов есть несколько типов фотосинтеза. Фотосинтезирующие прокариотные организмы представлены пурпурными и зелеными бактериями, а также большой группой широко распространенных в почвах цианобактерий, почвенные фотосинтезирующие эукариоты — водорослями. Особый тип фотофосфорилирования на мембране обнаружен у га- лобактерий — представителей архебактерий, обладающих красным пигментом родопсином, заменяющим хлорофилл. У эукариот и цианобактерий фотофосфорилирование сопровождается выделением
[
Пропионоеыв бактерии j
Группа
кишечной палочт
Оксапоацетат
Ын]
Ацетил
- СоА + АТР^/
|
нсоон
Ацетил
- СоА +
|н2|
|
СО;
|
ATP—f
Сукцииат
|
|
СО; |
Ацетоацвтил —СоА ATP-j-t"!
|
Ацетат | | Этаиоп |
V
СО,
| ; 1
j
Ацетоии
1

-ссь4
Пропмонат
|
Бутандиоп
Рис. 65. Типы брожений и их возбудители
кислорода, у остальных прокариот фотосинтез бескислородный (без выделения свободного кислорода).
Появление в атмосфере Земли кислорода в результате фотосинтеза привело к возникновению нового типа энергетического метаболизма, основанного на акцептировании электрона кислородом. Это так называемое окислительное фосфорилирование, в котором различают дыхание (перенос электрона на молекулярный кислород) и анаэробное дыхание (перенос на неорганические акцепторы — сульфаты, нитраты, карбонаты) (рис. 66).
Анаболические процессы, или конструктивный (строительный) метаболизм, связаны количественно главным образом с источниками углерода и азота, которые могут быть в органической и неорганической форме. Соотношение С : N в биомассе бактерий составляет 5:1, а в среде оптимальное соотношение должно быть 25:1, так как 1/5 часть углерода включается в вещества клеток, а 4/5 расходуется на энергетические нужды (это усредненные весьма ориентировочные расчеты). Источниками углерода служат С02 и(или) органические соединения. В качестве источников азота выступают белки, пептиды, аминокислоты, нитраты, аммонийные соединения и молекулярный азот. Кислород поступает в органическое вещество главным образом из С02, а водород — из воды.
Рис.
66.
Аэробное и анаэробное дыхание
Кроме источников основных элементов — органогенов, зольных элементов и микроэлементов многие микроорганизмы нуждаются в специфических веществах, которые лимитируют рост и поэтому называются факторами роста. Такими факторами у микроорганизмов могут быть витамины, аминокислоты, пуриновые и пиримидиновые основания. Независимые от органических факторов роста микроорганизмы называют прототрофами, а нуждающиеся в каком-либо ростовом факторе — ауксотрофами.
Специфические потребности микроорганизмов в источниках питания, факторах роста или микроэлементах делают их объектами, удобными для использования в качестве биологических индикаторов, например при определении потребности почвы в удобрениях, количественном анализе содержания витаминов или микроэлементов в почве. Впервые на свойства микроорганизмов как химических реактивов обратил внимание академик B.J1. Омелянский. В 1906 г. он написал статью «О применении бактериологического метода при химическом исследовании», в которой отметил две особенности микроорганизмов как биологических индикаторов: их специфичность и исключительную чувствительность.
Некоторые продукты метаболизма и особенно вторичные метаболиты являются ценными веществами. Это аминокислоты, лимонная кислота и другие органические кислоты, различные сахара, ферменты, витамины, антибиотики, которые составили целую эпоху в развитии микробиологии, в том числе и почвенной микробиологии. Микроорганизмы способны к «сверхсинтезу» некоторых веществ, которые образуются ими в избытке, выделяются в среду или аккумулируются в клетке. На этом свойстве основано промышленное использование микробов для получения разных целевых продуктов. Почва в таком случае выступает как неистощимый источник микроорганизмов, образующих разнообразные ценные вещества и в первую очередь антибиотики, витамины, стимуляторы роста растений.
Все процессы катаболизма и анаболизма протекают не самопроизвольно, а катализируются особыми ферментами. Клетка обладает способностью управлять скоростями отдельных реакций и тормозить или ускорять весь метаболический путь за счет наличия регуляторных механизмов.
Процессы анаболизма регулируются в клетке механизмом, который называется репрессией конечного продукта. Сигнал к прекращению синтеза белковых молекул в большинстве случаев поступает от конечных продуктов процесса. Он проявляется в том, что в цепи одного из метаболических путей есть фермент, имеющий регуляторный центр, который может связываться с конечным продуктом данного пути биосинтеза и изменять активность фермента (аллостерический фермент) (рис. 67).
Процессы катаболизма регулируются при помощи механизма катаболитной репрессии. Если в среде для роста имеется два субстрата, то бактерии в первую очередь используют тот из них, который обеспечивает более быстрый рост, а второй потребляют только по исчерпании первого — явление диа- уксии. В этом случае синтез ферментов для использования второго субстрата репрессируется первым катаболитом.
Ферменты делят на конститутивные, т.е. всегда синтезируемые организмом независимо от условий его роста, и индуцибельные, которые синтезируются клеткой только в присутствии соответствующего субстрата. В этом одна из
особенностей ферментных СИС- (отрицательный) ческий тем микроорганизмов. Другая фермент
особенность- образование Рис_ 67 Дллостерические ферменты экзоцеллюлярных (внеклеточных) ферментов, которые расщепляют сложные молекулы субстрата во внешней среде (целлюлоза, белки, лигнин, хитин, ли- пиды, пектин). При этом клетка может синтезировать много фермента и поддерживать высокую активность процесса превращения экзогенного субстрата. Результатом является переработка больших количеств разнообразных веществ микроорганизмами при малой их биомассе.
Ферменты характеризуются специфичностью действия и специфичностью субстрата. Последнее свойство используется при построении названий ферментов: к названию специфического для субстрата фермента прибавляется окончание «аза». Например, пектиназа, целлюлаза, липаза, фосфатаза, протеаза (от слова протеин — белок), амилаза (от слова амилоза — крахмал) и др.
0
.
ФЕРМЕНТ + ПРОДУКТ
Фермент
Фермент- субстратный комплекс
Эффектор
Аллостери-


усваивает субстрат, чем в отдельности каждая популяция, входящая в ее состав. В метабиотических цепях каждый последующий организм усваивает тот субстрат, который является конечным продуктом деятельности предшественника. В результате микроорганизмы оказываются способными утилизировать бесконечное множество различных соединений — природных и создаваемых человеком (ксенобиотиков). Наиболее стойкие природные соединения — лигнин и гумус — тоже подвергаются микробной деградации, хотя и медленнее, чем другие органические вещества. Неприродные синтезированные химиками органические соединения не всегда разлагаются микроорганизмами (многие полимеры и пестициды). Они опасны тем, что могут накапливаться в почве в угрожающих количествах.
Совокупность микроорганизмов, способных осуществлять одну и ту же общую физиологическую функцию в цепи превращения веществ, в микробиологии принято называть физиологической группой. Примерами могут служить физиологические группы микроорганизмов, участвующих в превращениях азота (нитри- фикаторы, аммонификаторы, денитрификаторы, азотфиксаторы) или серы (серные, тионовые, сульфатредуцирующие). Такое разделение микроорганизмов на физиологические группы закрепилось в почвенной микробиологии благодаря широкому внедрению принципа элективное™ С.Н. Виноградского: в специфических избирательных условиях может работать только определенный микроорганизм или группа организмов с четко обособленной функцией. В последнее десятилетие было получено много доказательств смены функций у одного и того же вида микроорганизма в зависимости от условий. Например, типичный азотфикси- рующий микроорганизм при наличии нитратов, органического вещества и в анаэробных условиях осуществляет денитрифика- цию и не фиксирует, а выделяет N2. При использовании белка он становится аммонификатором. Следовательно, понятие физиологической группы нельзя считать строго ограниченным определенным набором видов. Это подвижная группировка микроорганизмов, проявляющих в данный момент определенную функциональную активность при заданных условиях среды. Правильнее говорить не о физиологических группах, а о физиологических функциях микроорганизмов, которые могут проявляться по-разному в зависимости от условий, хотя есть и узкоспециализированные группы микробов, например, метаногены, автотроф- ные нитрификаторы, сульфатредукторы.
ЦИКЛ УГЛЕРОДА
Цикл углерода в почве — важный геохимический цикл, обеспечивающий стабильность углерод-кислородной системы Земли.
С круговоротом углерода связаны важнейшие процессы — фотосинтез и судьба почвенного органического вещества. От динамики этих процессов зависит состояние наземного покрова и атмосферы.
В круговороте углерода и кислорода наиболее четкое выражение находит взаимозависимость и связь всех живых существ на Земле. Благодаря участию в этом цикле микроорганизмов почвы как деструктивного звена происходит замыкание круговорота и совершается циклическое превращение веществ и энергии в биосфере. В этом главнейшая функция почвенных микроорганизмов.
С
А
Рис.
68.
Цикл углерода и кислорода

Углерод в составе биосферы представлен разнообразными формами — от простейших одноуглеродных соединений до сложнейших молекул лигнина. Рассмотрим разные механизмы синтеза углеродных соединений с участием микроорганизмов почвы и разложения органических веществ в ряду от наиболее простых к сложным и стабильным формам углеродных соединений.
Процессы связывания (фиксации) С02
Из углекислоты создается вся первичная органическая продукция. Содержание С02в атмосфере около 0,03%, в почвенном воздухе — на порядок больше. Биологическое связывание С02 происходит в процессах фотосинтеза, хемосинтеза и гетеротрофной фиксации. Фотосинтез в основном происходит в наземном слое биосферы, два последних процесса — преимущественно в почве.
Фотосинтез. Фиксация С02 в фотосинтезе происходит за счет световой энергии:
С02 + Н20 + hv -> (СН20) + 02,
где hv — световая энергия (рис. 69, 70).
На этот процесс в биосфере расходуется 0,1% энергии Солнца, попадающей на Землю. Запасенная в продуктах фотосинтеза энергия служит важным энергетическим источником для ряда процессов, происходящих на Земле. В результате фотосинтеза ежегодно создается 180 млрд т органического вещества, из них 120 млрдт на суше и 60 млрд т в океане и выделяется приблизительно 100 млрд т свободного 02. Фотосинтез осуществляется высшими растениями, водорослями и некоторыми бактериями. Первичную продукцию суши определил и точно рассчитал в 1864 г. немецкий химик Ю. Либих. В своих расчетах он основывался на продуктивности луга, находившегося около его лаборатории. В дальнейшем (1964— 1974 гг.) проводились исследования по Международной биологической программе. Исследователи определяли продуктивность в самых различных уголках земли: поднимались в горы, с вертолетов высаживались в недоступные районы тропических лесов и болот. Получили много ценных сведений о продуктивности различных экосистем, но в то же время установили, что цифры, рассчитанные Ю. Либихом, являются совершенно точными.
В фотосинтезе участвуют пигменты, содержащиеся в хлоро- пластах (у растений и водорослей) или в мембранных структурах бактерий — хлоросомах, тилакоидах, фикобилисомах.
Свет
Ш
би •eh
\
Фотособирающая
антенна
Реакционный
центр

КИСЛОРОДНЫЙ
ФОТОСИНТЕЗ растения;
Водоросли; цианобактерии
ДОНОРЫ
СО,
нг
орг.
в-ва
ОРГАНИЧЕСКОЕ
/\ -- ВЕЩЕСТВО < ~ „
БЕСКИСЛОРОДНЫЙ
ФОТОСИНТЕЗ пурпурные
серные и несерные бактерии ; зеленые
бактерии
ЭЛЕ КТРОНА
H2S '
Рис. 69. Схема фотосинтеза у растений, водорослей и бактерий
|Н]
|Н|
СОЛНЦЕ
л
и
другие органические соединении
ЭНЕРГИЯ
ИЗЛУЧЕНИЯ
СО,
ЭНТРОПИЯ
БИОХИМИЧЕСКАЯ ЭНЕРГИЯ И ТЕПЛО
н2о
Нг<)
-
Оргаиотрофиые
аэро5но дышащие организмы
Рис. 70. Фотосинтез и деструкция органических веществ
В основе фотосинтеза лежат окислительно-восстановительные процессы, в которых электроны переносятся от донора-восстановителя к акцептору. Восстановителем в растительном фотосинтезе выступает молекула воды, акцептором — С02. При этом
образуется органическое вещество и выделяется кислород. У бактерий донорами электрона могут быть сероводород, сера, водород, органические соединения. Кислород в этом процессе не выделяется (бескислородный фотосинтез) (рис. 69).
Фиксация С02у автотрофных организмов происходит за счет универсального механизма, известного как цикл Кальвина, или рибулозобифосфатный цикл. У зеленых бактерий имеется циклический механизм фиксации С02, отличный от цикла Кальвина. В его основе лежат реакции восстановительного карбоксилирова- ния органических кислот, получившие название цикла Арнона.
Фототрофные прокариоты составляют группу микроорганизмов с разным отношением к кислороду — от строгих анаэробов до аэробов, у которых кислород образуется внутриклеточно. Есть факультативные анаэробы, аэротолерантные формы и микро- аэрофилы.
Из фототрофных прокариот в почвах обитают цианобакте- рии, галобактерии (на солончаках) и пурпурные несерные бактерии в болотах и на рисовых полях. Некоторые фототрофные прокариоты развиваются на мхах, папоротниках, высших растениях, они также вступают в более тесные ассоциации с другими организмами, например цианобактерии с грибами образуют некоторые лишайники.
Хемосинтез. Фиксация С02 в хемосинтезе происходит за счет энергии окисления внешнего неорганического донора электрона. Этот процесс впервые был описан С.Н. Виноградским в конце прошлого века и назван аноргоксидацией. Хемоавтотрофы известны только среди бактерий (прокариот). К хемосинтезиру- ющим организмам относятся нитрификаторы, серобактерии, железобактерии, водородные бактерии, карбоксидобактерии. Они называются так по субстратам окисления, которыми могут быть NH3, NO2", H2S, S, Fe2+, H2, CO. Некоторые виды — облигатные хемолитоавтотрофы, другие — факультативные. К последним относятся карбоксидобактерии и водородные бактерии. В отсутствие органических веществ они используют энергию окисления водорода или окиси углерода для фиксации С02, а при наличии органических соединений переходят на гетеротрофный тип питания, т.е. они являются миксотрофами.
Гетеротрофная фиксация С02. Помимо фото- и хемосинтеза, где углекислота служит единственным источником углерода, она может ассимилироваться гетеротрофно, путем присоединения к готовым углеродным цепям. К гетеротрофному связыванию углекислоты способны и такие фотоавтотрофы как высшие растения, а также животные. Акцепторами С02 могут быть различные органические кислоты, чаще всего, пировиноградная (реакция Вуда-Веркмана):
СНзОСООН + С02 СООНСН2СОСООН,
хотя всего известно до 10-12 способов включения С02 при гетеротрофной фиксации.
До 3-10% углерода биомассы гетеротрофных организмов может быть построено из С02, как это было показано в опытах при использовании меченой |4С02.
Другие пути превращения одноуглеродных соединений
Образование метана биологическим путем из С02 и других одноуглеродных источников — метановое брожение — происходит в болотах, торфяниках, иловых отложениях озер, метантен- ках, рубце жвачных животных, кишечном тракте человека. В почве этот процесс протекает там, где возникают анаэробные условия, и образуется водород в первичных процессах анаэробного превращения органических веществ.
Метанобразующие бактерии (метаногены) — вторичные анаэробы, так как они перерабатывают продукты, поступающие от других анаэробов. Метан образуется за счет восстановления С02 водородом по следующей схеме:
С02 + 4Н2 = СН4 + 2Н20.
Восстановлению могут подвергаться также СО, муравьиная кислота и метанол.
В газовой атмосфере торфяников метан часто превышает 30%. Группа метанобразующих бактерий гетерогенна. Среди них есть кокковидные, палочковидные, сарциноподобные организмы и спириллы. Все метаногенные бактерии относятся к архебакте- риям, их родовые названия начинаются со слова methan (Methano- coccus, Methanosarcina, Methanobacterium, Methanospirillum). Метанобразующие бактерии строгие анаэробы. Кислород убивает их, с чем сопряжены большие трудности при работе с ними в лаборатории. Необходимо применять специальные методы, которые были разработаны в последние 20 лет, и было проведено подробное их изучение. В ряде анаэробных экосистем основным субстратом для образования метана служит ацетат. Метанобразующие бактерии составляют последнее звено анаэробной пищевой цепи, в начале которой находятся полисахариды (целлюлоза, крахмал, хитин), липиды, белки. В этой цепи участвуют: 1) бактерии, сбраживающие целлюлозу до сукцината, пропионата, бутирата, лактата, ацетата, спиртов, С02 и Н2; 2) ацетогенные бактерии, сбраживающие эти первичные продукты до ацетата, формиата, С02 и Н2 (рис. 71).
Рис.
71.
Аэробное и анаэробное разложение
растительной биомассы
Метанобразующие бактерии находятся в симбиотических (му- туалистических) взаимоотношениях с бактериями, образующими газообразный водород. Первые не могут развиваться без водорода, а вторые сами себя отравляют избытком образуемого ими водорода. При совместном развитии этих двух групп бактерий свободный водород практически не обнаруживается, так как он моментально используется метанотрофами.
Метанобразующие бактерии активируют водород и осуществляют его окисление, сопряженное с восстановлением С02. Клеточное вещество они также строят из С02, т.е. являются хемоав- тотрофами. Они осуществляют карбонатное дыхание (в качестве источника С02 используют СаС03). Ферменты метанобразующих бактерий отличаются большой специфичностью. Образование метана может происходить из ацетата или С02
Автотрофная фиксация С02 у метанобразующих бактерий происходит без участия рибулозобифосфатного цикла.
В последнее десятилетие выделение метана в атмосферу увеличивается на 1% в год, что связано с воздействием на природу антропогенного фактора (утечки при газодобыче, рисовые поля, скот). Однако в атмосфере он фотохимическим путем довольно быстро окисляется до С02. В ряде случаев почвы могут поглощать метан. Вопрос о том, что же почвы земного шара больше выделяют или поглощают метан, остается открытым.
Бытовые органические отбросы в южных странах используются для получения метана в специальных метантенках (биогаз), который используется в домашнем хозяйстве. Метантенки бывают на один или несколько домов. В северных странах при проведении микробиологического процесса почти весь метан приходится использовать для подогрева метантенка.
Метан, помимо биологического пути, образуется и в результате геохимических процессов. Он составляет основную часть природных углеводородных газов.
Окисление метана (метанотрофы) — биологический процесс, активно протекающий в районах метаногенеза — в местах газовых и нефтяных месторождений, в переувлажненных почвах, в поверхностной пленке болот. Метанокисляющие бактерии перехватывают метан, который образуется в анаэробной зоне, и окисляют его до С02 через метанол, формальдегид и муравьиную кислоту:
СН4 СН3ОН -> СНОН -> НСООН С02.
Этот путь диссимиляции (катаболизма) осуществляется с участием ферментов цепи переноса электронов. Ассимиляция углерода метана происходит на уровне формальдегида несколькими путями, отличными от автотрофной фиксации С02 в цикле Кальвина. Микроорганизмы, окисляющие метан, получили название метанотрофов. Среди них есть облигатные формы, использующие в качестве углеродного источника питания метан или его производные (метанол и метиламины) и факультативные мети- лотрофы, ассимилирующие как одноуглеродные соединения (метанол, формальдегид, метиламин, но не метан), так и другие органические вещества. Облигатные метанотрофы — группа ме- танокисляющих бактерий семейства Methylomonadaceae, которые относят к родам Methylomonas, Methylobacillus, Methylococcus, Methylosinus и др. Хотя формально их следует считать гетеротро- фами (метан — органическое вещество), но по многим морфологическим и физиологическим признакам они близки к таким литоавтотрофам, как нитрифицирующие бактерии семейства Nitrobacteriaceae. Процесс окисления метана сопровождается у них окислением аммиака до нитритов, они имеют хорошо развитую систему внутриклеточных цитоплазматических мембран, пути метаболизма метанокисляющих бактерий сходны с таковыми ав- тотрофных нитратных бактерий.
Среди факультативных метилотрофов известны не только прокариоты, но и представители дрожжей.
Метанокисляющие бактерии, вовлекая в биологический круговорот одноуглеродные соединения группы метана, вносят значительный вклад в глобальный цикл углерода, замыкая трофические цепи в специфических нишах биосферы. Они представляют интерес и в практическом отношении как продуценты белка из дешевого сырья, а также как средство борьбы с метаном в угольных шахтах для предотвращения взрывов.
Окисление окиси углерода микроорганизмами — процесс, поддерживающий природное равновесие этого газа в биосфере; СО образуется техногенным путем в результате неполного сгорания разных видов топлива. Существуют и природные источники угарного газа — извержения вулканов и биохимические реакции, приводящие к образованию СО при распаде порфиринов, флавано- идов, серина и некоторых других органических веществ. Оксид углерода связывают животные, растения и микроорганизмы. Это обусловлено его взаимодействием с гемоглобином, цитохромок- сидазой и рядом других ферментов. В результате происходит нарушение функций организмов, для большинства из которых оксид углерода даже в небольшой концентрации очень токсичен. Повышение его содержания в помещении может привести к гибели человека и животных (отравление угарным газом). Чувствительность к СО проявляют и многие микроорганизмы, однако некоторые используют его в качестве источника энергии и углерода для построения своего тела. Большинство этих микроорганизмов помимо СО окисляют молекулярный водород.
Бактерии, использующие СО в аэробных условиях, называются карбоксидобактериями, или карбокситрофами.
Удаление окиси углерода из атмосферы происходит в результате ее окисления кислородом воздуха и окисления почвенными микроорганизмами. Ежегодно почва поглощает 4 х 1014 г СО, что лишь немногим меньше, чем образуется при сгорании топлива. Таким образом, именно почва поддерживает равновесное состояние окиси углерода в атмосфере.
В окислении окиси углерода участвуют микроорганизмы группы карбоксидобактерий, которые не обладают морфофизиоло- гическим единством и относятся к разным родовым и видовым таксонам (Alcaligenes, Acinetobacter, Pseudomonas, Achromobacter, Comamonas, Arthrobacter, Bacillus, Streptomyces, Hydrogenophaga, Seliberia). Карбоксидобактерии — автотрофы. Образующаяся в результате окисления СО углекислота фиксируется в процессах анаболизма автотрофно через цикл Кальвина, а не метилотроф- но. Но автотрофия карбоксидобактерий не облигатная. Они способны и к органотрофному росту. Однако высокие концентрации органических веществ подавляют их развитие, на основании чего их относят к олиготрофной группировке почв. Помимо карбоксидобактерий СО окисляют неспецифические микроорганизмы в процессах кометаболизма. Возможно, по масштабам этот процесс значительно превышает окисление СО карбоксил оба к- териями.