

Концепция почвы как множества сред обитания микроорганизмов
С микробиологических позиций почва представляет собой крайне гетерогенную среду и не может рассматриваться как единая однородная среда обитания. Благодаря своей структурированности и микрозональности она должна рассматриваться, как набор различных микро- и мезосред, в каждой из которых создаются различные и часто прямо противоположные условия для развития отдельных групп микроорганизмов. Множество таких микросред может находиться в каждом грамме почвы. Микро- и мезозоны разделены в пространстве и времени. Микроорганизмы — это как раз такие организмы, которые адаптированы к развитию в микрозонах. Микроскопические размеры дают им возможность осваивать микросреду. Способность быстро размножаться и быстро переходить к покою или крайне замедленному метаболизму дает им возможность за короткий срок освоить микрозону и выжить при исчерпании запасов питания. Микрозоны могут быть очень небольшими и занимать всего несколько десятков или сотен кубических микрометров. При этом в микрозоне часто развивается одна микроколония, состоящая из нескольких десятков клеток одного вида. Иногда такие зоны имеют значительные размеры, например кусок разлагающихся растительных остатков. Они могут иметь большую протяженность при небольшой толщине, например, поверхность однородного участка корня (ризоплана).
Микрозональность основывается на локальном поступлении органических остатков и корневых выделений, а также на микрозональности распределения физико-химических условий (окислительно-восстановительного потенциала, рН, концентрации элементов питания и т.д.), минералогических факторов.
Несмотря на огромное количество микроорганизмов, содержащихся в почве (миллиарды на 1 г), оказывается, что клетки, как правило, собраны в микроколонии, разделенные пустыми пространствами, которые по площади в сотни и тысячи раз превосходят пространства, занятые микроорганизмами. Таким образом, микроколонии, состоящие из клеток одного или нескольких видов, могут развиваться сравнительно изолированно. Отсюда следует важный вывод о том, что в почве часто развиваются чистые микрокультуры микроорганизмов. Это положение было подтверждено экспериментально при изучении разных почв с помощью люминесцентной микроскопии. Пространства, разделяющие микроколонии, настолько велики по сравнению с размерами самих колоний, что трудно ожидать их тесного взаимовлияния. Следовательно, изучение чистых культур имеет более прямое отношение к почвенной микробиологии, чем считалось ранее. Основной вопрос, который в настоящее время нуждается в разрешении: насколько развитие в микрокультуре (микроколония) соответствует развитию в макрокультуре на чашках Петри (макроколония). Если такое соответствие можно признать достаточно полным, то данные, полученные в лабораторных условиях на чистых культурах, можно переносить на почвенные условия. Если развитие в тонких пленках воды, в капиллярах, а также просто в микрокультурах отличается (например, есть сведения, что микрокультуры клостридиума не образуют ботулина), то необходимо разрабатывать методы для изучения микрокультур. Метод чистых культур дает возможность изучать только генетические возможности культуры, а конкретное проявление будет зависеть от экологических условий.
В связи с концепцией микрозональности становятся понятными многие аспекты строения и функционирования комплекса почвенных микроорганизмов. Один из важных вопросов состоит в связи микрозональности с широким географическим распространением каждого вида микроорганизма. Дисперсия микроорганизмов, особенно воздушная, обеспечивает попадание всех почвенных микроорганизмов во все почвы. Если учесть, что среда обитания микроорганизмов является микросредой, которая может быть прерывистой во времени, то понятно, что все или почти все почвенные микроорганизмы могут выживать в некоторых микрозонах любых почв («все есть всюду» — правило Бейе- ринка). Однако в глобальных процессах круговорота веществ существенное экологическое значение будут иметь не все почвенные микроорганизмы, а только те, которые многочисленны и проявляют активную жизнедеятельность (пул высокого уровня). При проведении экологических исследований в большинстве случаев нет необходимости устанавливать все существующее в почве разнообразие микроорганизмов, а достаточно ограничиться установлением доминирующих в важных экологических процессах форм. Таким образом, критерии доминирования приобретают первостепенное значение для почвенной микробиологии, причем имеется в виду не только и не столько численное доминирование, а доминирование в проведении процессов. В одних случаях это могут быть такие глобальные процессы, как разложение целлюлозы или азотфиксация, а в других — синтез определенного витамина или определенной аминокислоты. К сожалению, в настоящее время в большинстве случаев почвоведы-микробиологи могут учитывать только численность определенных видов или даже родов микроорганизмов в почве, да и то с большим трудом и большими погрешностями. Интенсивность процессов, вызываемых определенным видом, почти не поддается определению. Комбинация методов иммунолюминесценции и авторадиографии позволяет не только видеть клетки определенного штамма микроорганизма в нестерильной почве, но и оценить активность каждой обнаруженной при микроскопии клетки. Но такие методы сложны и трудоемки. В связи с этим часто приходится довольствоваться условными критериями численности. Для бактерий может бьггь принят условный критерий, который гласит, что существенное экологическое значение бактерии имеют в том случае, если число их клеток не менее 1 млн на 1 мл или 1 г субстрата.
Поиски более редких бактерий могут иметь значение для индикации почв и других целей, но мало дают для оценки процессов, происходящих на данном этапе развития экосистемы. Для грибов, дрожжей, водорослей и других необходимо разрабатывать свои критерии экологической значимости, учитывая количество микроорганизмов, их размеры и интенсивность метаболизма. Поскольку эти организмы по массе приблизительно в 100 раз крупнее, чем бактерии, то для них может быть принят критерий 10 тыс. на 1 г.
Физиологически активные вещества в почве (антибиотики, токсины, витамины, аминокислоты, стимуляторы и ингибиторы роста растений) важны для развития микроорганизмов в микрозонах. В то же время их глобальная регулирующая роль в масштабах всей почвы, хотя и признавалась некоторыми авторами, нуждается в дальнейшем экспериментальном подтверждении или опровержении. Первое место в качестве таких регуляторов, очевидно, могут занимать газообразные вещества, так как они наиболее быстро диффундируют и могут передавать информацию, являясь «средовыми гормонами». Действие физиологически активных веществ, в том числе газов или летучих органических веществ, в качестве «средовых гормонов» легко обнаружить в искусственных условиях, когда в почву вносятся большие количества органического вещества, и она перестает функционировать как система микросред и действует как единая система, т.е. переходит в другой тип микробных экосистем. Такое состояние возможно для больших частей естественной почвы на небольших отрезках времени, но оно не характерно для всей массы почвы. В почве имеются определенные зоны, функционирующие по другим законам. Прежде всего это зона ризопланы и в какой-то мере ризосферы, пищеварительный тракт беспозвоночных животных, а также подстилка в лесу, степной войлок, разлагающийся торф и др. В этих случаях можно ожидать и искать большую организованность процессов, чем в других частях почвы.
Важным открытием, которое должно использоваться в экологии почвенных микроорганизмов, является кометаболизм. Установлено, что микроорганизмы могут разрушать некоторые труднодоступные вещества, в том числе и неприродные органические вещества. Принципы строения и функционирования комплекса почвенных микроорганизмов должны широко использоваться при разработке критериев целостности и повреждения этого комплекса в связи с применением удобрений, пестицидов, загрязнением почвы, а также при решении вопросов о целесообразности внесения в почву тех или иных «полезных» микроорганизмов или регулирования комплекса почвенных микроорганизмов путем изменения среды обитания.
ЭКОЛОГИЯ КЛЕТКИ
Аутэкология организма — это важный и хорошо разработанный раздел современной экологии животных и растений. В отношении почвенных микроорганизмов он почти не разрабатывался, так как считали, что клетки настолько малы, что здесь трудно получить ценные сведения. Вся физиология и биохимия разрабатываются на популяционном уровне (в отличие от растений и животных). Однако аутэкологии микроорганизмов следует уделять больше внимания. Изучение величины и морфологии клетки дает много ценного.
Организм и среда неразделимы. Эколог говорит: «Скажите, какова среда, и я отвечу, какие организмы здесь обитают. Скажите, каковы организмы, и я отвечу на вопрос об особенностях среды». Если известно кое-что о природе окружающих условий, то можно предсказать экологические особенности организмов, можем найти их. Напротив, если известно что-то об организмах, которых нашли в данном местообитании, тогда можно предсказать кое-что о природе окружающих условий. При проведении аутэкологических исследований исследователь должен постоянно переходить от опытов с лабораторными средами к опытам с почвой, используя все наблюдения как дополняющие друг друга.
Формы микроорганизмов разнообразны, что имеет большой экологический смысл (рис. 104). Форма фаготрофных организмов, например амеб, определяется их потребностью поглощать клетки бактерий или частицы органического вещества. Однако подавляющее большинство микроорганизмов поглощает только молекулы, причем в основном мелкие молекулы, но не полимеры. Форма палочек, спирилл и нитей по сравнению с шаром (кокки) увеличивает поверхность клеточной мембраны. В бедных средах больше этих форм, чем кокков. Это правило часто не выполняется для почв, однако, здесь кокковидную форму обычно имеют покоящиеся клетки. Сферические формы более устойчивы к высушиванию, вероятно, из-за более равномерного давления на клетку во время высушивания. Кокки и дрожжи являются наиболее обыкновенными аспорогенными микроорганизмами воздуха.
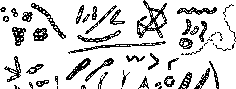
Рис.
104.
Морфологическое различие типов
бактериальных клеток
Олиготрофные бактерии в почве имеют различные выросты, стебельки, простеки и другие, что позволяет им увеличить площадь поверхности и собирать питание из разбавленных растворов. Строение клетка-стебелек-клетка позволяет видам, откладывающим на своей поверхности чехол из железа и марганца и образующим в почве ортштейны, выбраться из чехла и продолжать рост на поверхности железо-марганцевых конкреций (сравни с кораллами). Некоторые почвенные бактерии на богатой питательной среде имеют форму шара, но на бедной среде приобретают форму звезды (Stella humosa), т.е. резко увеличивают площадь поверхности.
Особое значение для жизни в почве имеет мицелиальное строение (грибы, актиномицеты). Нитчатое строение с верхушечным ростом в почве как в гетерогенной микрозональной среде дает такому микроорганизму явное преимущество. Он преодолевает микрозоны без источников питания и ищет благоприятные условия. Мицелиальные микроорганизмы — это типичные почвенные организмы (в отличие от водных). В почве грибы победили бактерии и стали главными потребителями растительных остатков, хотя в прежние геологические эпохи в воде в цианобактери- альных матах, которые господствовали на земле, основными деструкторами были бактерии. Толщина мицелия актиномицетов в 5-10 раз тоньше мицелия грибов, они медленно растут и им труднее преодолевать большие пространства. Однако по биомассе в почве они составляют 25% общей биомассы прокариот. В их прогрессе большое значение имеет то, что они обладают мицелиаль- ным ростом, являются активными гидролитиками и образуют разнообразные антибиотики (стрептомицин, тетрациклины, ле- вомицетин и многие другие).
Размеры микроорганизмов имеют важнейшее экологическое значение и по диаметру варьируют в десятки тысяч раз: вирус 200 А, микоплазма 1000 А, бактерия 10 тыс. А (1 мкм), толщина гиф фибов 100 тыс. А (10 мкм), амеба 1 млн А (100 мкм). Одни из них благодаря фавитационным силам подвергаются быстрому осаждению, другие из-за броуновского движения вообще не осаждаются. Для осаждения большую роль ифает подвижность клеток. Величина клеток имеет большое значение для скорости обменных процессов. В экологии существует общий закон: чем крупнее организм, тем медленнее он размножается. До сих пор неясно, насколько этот закон применим к микроорганизмам, видимо, если он и применим, то только частично. Максимальная удельная скорость роста микроорганизмов запрограммирована генетически.
Ригидность клеточной стенки микробов очень важна для поддержания внутри клетки повышенного осмотического давления. Клетки экстремальных галофилов (архебактерий) находятся в осмотическом равновесии с окружающей средой. Их клеточные стенки имеют особое строение и стабилизируются ионами калия и натрия.
Большое значение для экологии имеют поверхностные структуры. Гидрофильность или гидрофобность клеточной поверхности определяет местоположение клетки в жидкой среде. Гидрофобные клетки образуют на поверхности пленку, они располагаются на гидрофобных участках поверхности почвенных частиц, а гидрофильные — в водной толще или на дне. Они будут адгезиро- ваться на гидрофильных почвенных частицах. Важен положительный или отрицательный заряд клетки. Правда, в большинстве случаев он мозаичный. На поверхности клеток бактерий часто находятся фимбрии и пили, которые служат для прикрепления или переноса генетического материала. В некоторых случаях адгезия клеток определяется специфическими химическими взаимодействиями адгезина и рецептора по принципу ключ-замок.
Клеточные капсулы предохраняют клетки от действия ферментов и обусловливают устойчивость к неблагоприятным факторам. Капсулы часто служат убежищем для микробов-спутников.
Клетки некоторых бактерий имеют внутриклеточные газовые вакуоли, а другие — внеклеточные газовые баллоны. Они важны для газообмена и для изменения положения клеток в воде, капиллярах и водных пленках.
Особый интерес в последнее десятилетие вызывают магнито- бактерии, содержащие в своих клетках магнитосомы с мельчайшими магнитиками (до 100 частиц на клетку). Они дают ориентацию клеткам в магнитном поле Земли. В Северном полушарии клетки должны двигаться к Северному полюсу. Экологическое объяснение этому явлению трудно найти. Однако по линиям магнитного поля клетки должны двигаться не просто на север, но и на дно водоемов, чтобы оказаться в толще ила. Это уже может иметь разумное экологическое объяснение. Магнитобак- терии в большом количестве содержатся и в почве. Зачем им нужны магнитные свойства, пока трудно объяснить.
Движение с помощью жгутиков или скользящее движение помогает клеткам выбирать наиболее благоприятную зону, а также способствует их расселению.
Эндоспоры бацилл, клостридий и стадии глубокого анабиоза у других родов бактерий представляют большой интерес для аут- экологии как стадии жизни, в которых не выявляются метаболические процессы. Эти формы устойчивы к экстремальным условиям и отличаются долговечностью.
Особо изучаются половые споры грибов, которые характеризуются специфическими свойствами. Конидии грибов и актиномицетов обладают существенным метаболизмом, но также служат для сохранения вида. Грибы очень долго могут сохранять жизнеспособность в сухом состоянии. Экспериментально показано, что это могут быть тысячи лет, например, в гробницах фараонов, а в замороженном состоянии в вечной мерзлоте — миллионы лет. Вероятно, фибы, как и бактерии, имеют структуры, сохраняющиеся в состоянии глубокого покоя очень долго даже при физиологически благоприятных условиях (влажность, температура, питание), но здесь нужны экспериментальные доказательства.
Имеется еще множество свойств клеток, которые должна рассматривать аутэкология.
ЭКОЛОГИЯ ПОПУЛЯЦИЙ
Экология популяций чистых культур микроорганизмов изучена наиболее досконально по сравнению с животными и растениями. Кинетика роста микробных популяций выражена в математических формулах, установлены многие важные закономерности роста. Однако изучение микробных популяций непосредственно в почве не проводилось, так как не было методов для проведения таких исследований, но в последнее время такие методы появились.
Генетическая маркировка. В почву вносится штамм микроорганизма, маркированный по какому-либо свойству, например, антибиотикоустойчивости. Он вносится в почву и затем следят за его судьбой. Для определения численности внесенного микроорганизма на разных этапах развития из почвы делаются высевы на питательную среду с антибиотиком, являющуюся элективной для данного штамма. Колонии дают только клетки внесенного штамма, а собственно почвенные микроорганизмы не растут. Генетическая маркировка проводится также при введении генов, кодирующих образование фермента, расщепляющего бесцветное вещество с освобождением его части, обладающей окраской. Тогда колонии внесенного штамма в отличие от всех остальных оказываются окрашенными, их легко отличить.
Микроскопическая иммунолюминесценция (метод флуоресцирующих антител). С помощью люминесцентной микроскопии изучается серологическая реакция между антителами и антигенами, причем до реакции антитела соединяются с флуорохромами.
Метод основан на способности флуорохромов вступать в связь с антителами без ущерба для присущей антителу специфичности в связывании антигена. Для разных объектов эта реакция может быть штаммо-, видо- или родоспецифичной. Для клубеньковых бактерий она использовалась наиболее часто и является штам- моспецифичной.
3. Использование штаммов или видов легко определяемых по своим культуральным свойствам. Это либо специфический вид колоний (бациллы, многие актиномицеты и грибы), либо образование пигмента.
Для изучения динамики популяции был использован актино- мицет, образующий люминесцирующий антибиотик гелиомицин. Благодаря наличию антибиотика колонии актиномицета при освещении их ультрафиолетовыми лучами приобретают характерное яркое оранжевое свечение и тогда их легко отличить среди других микроорганизмов, вырастающих на питательной среде. Полевые опыты, продолжавшиеся в течение года, показали (рис. 105),
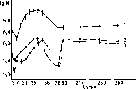
Рис. 105. Динамика популяции Streptomyces olivocinereus и аутохтонных актиномицетов в дерново-подзолистой почве (полевой опыт):
/ — аутохтонные актиномицеты; 2 — Str. olivocinereus (внесен мицелий); 3— Str. olivocinereus (внесены споры); N — численность колоний
вития актиномицета во времени. Резкие колебания численности отмечаются на протяжении первых трех месяцев после внесения. Затем популяционная численность стабилизируется на уровне 106 клеток на 1 г и остается постоянной в течение девяти месяцев. Внесенный актиномицет вписывается в общий актиноми- цетный комплекс почвы и в общих чертах повторяет динамику его развития. Форма внесения актиномицета (споры или мицелий) не оказывает решающего влияния на общую картину динамики.
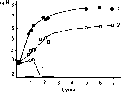
Динамика популяции этого актиномицета зависит от особенностей типа почвы (рис. 106). В среднеазиатском сероземе, из которого этот актиномицет был выделен, после падения численности он
дает резким подъем и затем ста- ig n билизируется на очень высо- 9 ком уровне 107, а в кислом красноземе он погибает. 8
В ризосфере пшеницы 7 одни бактерии быстро погибают, другие успешно размно- 6 жаются (рис. 107). 5
7
14
Рис.
106.
Динамика популяций
Str. olivocinereus
в
различных типах почв
Рис.
107.
Динамика популяций
Curtobac- terium (/), Mycoplana
sp.
(2)
и
Pseudo- monas
sp.
(J)
в
ризосфере пшеницы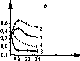
Сутки
Сутки
Рис. 108. Динамика популяций клубеньковых бактерий в различных типах почв и в разных условиях: