

Доминирующие роды бактерий в разных ярусах лесного биогеоценоза
Ярус |
Горизонт, субстрат |
Род |
Функционально- трофические группы |
Надземный |
Кора ели, шишки, хвоя, листья деревьев и кустарников |
Aquaspirillum Micrococcus Rhodococcus Flavobacterium |
Эккрисотрофы, ко- пиотрофы |
Травы, злаки, осоки |
Erwinia Methylobacterium Clavibacter- Curto- bacterium |
||
Наземный |
01 (L) |
Cellulomonas Cytophaga Myxobacterales |
Подстилочные са- протрофы |
02+03 (F+H) |
Bacillus Streptomyces Rhodococcus Cellulomonas Myxobacterales |
Гидролитики |
|
Почвенный |
А1 |
Streptomyces Bacillus Nocardia Rhodococcus |
Педобионты-сапро- трофы, гидролитики |
Группа почкующихся и стебельковых олиготрофных бактерий |
Олиготрофы |
трофическое окружение центрального ядра, т.е. растения-продуцента, и принадлежат к классу протеобактерий.
По кинетическим параметрам роста бактерии филлопланы относятся к популяциям, характеризующимся низкой инерционностью, высокими скоростями роста, но и быстрым отмиранием клеток при прекращении поступления питательных веществ, слабой конкурентной способностью, что позволяет отнести их к г-стратегам.
Протеобактерий являются доминирующей группировкой и в подстилке, особенно в ее верхнем слое L, куда они попадают с опадом. В этом ярусе они переходят с эккрисотрофного на сапро- трофное питание, используя водорастворимые мономеры, содержащиеся в растительных остатках, и начиная тем самым процесс деструкции растительных остатков. Набор таксонов, входящих в класс протеобактерий, в подстилке (в ее верхнем слое) тот же, что и в филлоплане.
В нижних слоях подстилки (F+H) появляются спорообразующие бактерии (Bacillus), цитофаги и миксобактерии; увеличивается доля стрептомицетов и целлюломонад. Все они принадлежат к функциональной группе гидролитиков, а по типу экологической стратегии являются L- и К-стратегами. Известно, что миксобактерии, цитофаги и целлюломонады способны разрушать целлюлозу, бациллы — крахмал и пектин, стрептомицеты — хитин. Таким образом, в нижних слоях подстилки происходит более глубокая переработка растительных остатков, в результате которой расщепляются сложные полимеры. Следует помнить, что основную функцию по деструкции растительных остатков в лесных биогеоценозах играют фибы. Однако нахождение бактерий в значительных количествах в тех биотопах, где идет активная переработка растительного опада, позволяет говорить об активном участии и бактериального компонента биоты в этих процессах. В целом подстилка является тем биотопом, где сосредоточено максимальное разнообразие бактерий, осуществляющих различные функции. Здесь есть и копиотрофы, и гидролитики, и олиготрофы, представленные разными родами. Подстилку можно рассматривать как экотон — переход между четко различающимися сообществами, богатыми видами, так как он включает представителей обоих сообществ (фитопланы и почвы) и формы, свойственные самому экотону (подстилке).
При переходе от наземного к почвенному ярусу уменьшается не только численность бактерий, но и их таксономическое разнообразие. В сапротрофном комплексе бактерий остаются главным образом стрептомицеты (их доля увеличивается), родокок- ки, бациллы, появляются нокардии. Типичными обитателями почвенного яруса, не обнаруживаемыми в других субстратах, являются такие олиготрофы, как Pedomicrobium, Metallogenium, Seliberia, Gallionella. Эти организмы в профиле почв образуют мощные разрастания и создают обогащенные окисным железом и марганцем очаги и прослойки (ортштейны, ортзанды). Часто они развиваются на поверхности фибных гиф. Однако все перечисленные выше организмы — олиготрофы — не выделяются обычными методами посева из разведений на общепринятые питательные среды; требуются специальные методы для их выделения из почв. Это же относится к другой фуппе специфических олиготрофных бактерий — простекобактерий, включающих уникальные по морфологии роды. Все эти бактерии способны расти при низких концентрациях питательных веществ, усваивая их из рассеянного состояния, т.е. принадлежат к «микрофлоре рассеяния». Они характеризуются экономным обменом, приспособлены к условиям конкуренции с другими организмами, обнаруживаются на поздних стадиях сукцессии, что позволяет отнести их к К-стратегам. К этой же экологической группе относятся и представители коринеподобных бактерий — артробактер и родокок- ки. Однако артробактер — типичный обитатель большинства почв — не обнаруживается в кислых лесных подзолистых почвах.
Таким образом, по вертикальной структуре лесного биогеоценоза меняется как таксономическая структура бактериального комплекса, так и принадлежность к определенным функциональным группам и типам экологический стратегий (см. табл. 13). Смена таксономического состава бактериоценоза происходит путем последовательного обогащения каждого из горизонтов новыми формами с сохранением таксонов, характерных для вышележащих горизонтов. Так, в подстилке при сохранении бактерий, типичных для филлопланы, появляются бациллы и стрептомицеты, которые становятся доминирующими в почвенном ярусе. По вертикали лесного биогеоценоза наблюдается природный континуум бактериальных сообществ, создаваемый теми таксонами, которые ассоциированы с живыми растениями или растительными остатками и практически обнаруживаются во всех ярусах. Это эккрисотрофы, переходящие в сапротрофный ярус, представленные спириллами, флавобактериями, цитофагами, микрококками. Они же обнаруживаются в почвенном ярусе в ризоплане растений и кишечнике почвенных беспозвоночных. На их фоне происходит смена групп бактерий, осуществляющих поэтапно процессы разложения растительных остатков. Доминирующие на определенной стадии сукцессии бактериальные таксоны (ярусные доминанты) позволяют говорить о стратификации и дискретности бактериальных сообществ в вертикальной структуре биогеоценоза.
Смена трофических групп по вертикали биогеоценоза отражает процесс сукцессии при разложении растительных остатков от псевдомонад и бацилл к актином и цетам, нокардиям и обли- гатным олиготрофам — факт, установленный в модельных опытах. В соответствии со сменой трофических групп меняется и тип стратегии. Следует отметить, однако, что подобное разделение условно и отражает лишь тенденцию к смене эккрисотро- фов r-стратегов на растениях — олиготрофами К-стратегами — в почвенных горизонтах. При этом в таком сложном биокосном теле, как почва, условно выделяется лишь качественно новый по сравнению с вышележащими биогеоценотическими горизонтами субстрат — минеральный компонент почв. Именно в этом субстрате способны обитать олиготрофные бактерии.
Однако это не означает, что каждому из компонентов биогеоценоза должна строго соответствовать только одна группа микроорганизмов со специфическими функциями и типом стратегий. Так в подстилке, где зарегистрированы максимальная численность и максимальное разнообразие бактерий эпифитносапротрофного комплекса, одновременно функционируют и копиотрофы, и гид- ролитики, и олиготрофы. В почвенном ярусе сосредоточены не только олиготрофы: копиотрофы обитают в ризоплане растений, там же постоянно обнаруживаются гидролитики, представленные как бациллами, так и бактериями актиномицетной линии. Разные группировки микроорганизмов особенно четко раскрываются в процессе микробной сукцессии.
Разнообразие экологических и трофических группировок бактерий во всех ярусах лесных биогеоценозов обеспечивает осуществление ими разнообразных функций.
Актиномицеты. В лесных экосистемах наблюдается следующая закономерность: при переходе от надземного яруса (филло- сфера растений) к наземному (подстилка) наблюдается резкое увеличение численности актиномицетов Если в надземном ярусе численность актиномицетов не превышает сотен тысяч КОЕ/г субстрата, то в подстилке, особенно в слое F+H, верхний предел численности актиномицетов достигает миллиона КОЕ/г почвы.
Известно и экспериментально доказано участие актиномицетов в минерализации растительных остатков, разложении гумусовых соединений. Неоднократно подтверждалось участие актиномицетов в деградации труднодоступных растительных полимеров, прежде всего целлюлозы. Внесение растительных остатков в почву часто сопровождается увеличением количества актиномицетов, учитываемых методом посева. Деструкция органического вещества начинается в лесных биогеоценозах с подстилки. Следовательно, вполне закономерно ожидать заселения этого местообитания представителями порядка Actinomycetales.
При переходе от наземного к почвенному ярусу в хвойных и лиственных лесах наблюдается либо снижение численности актиномицетов (там, где почвы отличаются низким содержанием гумуса, низкими значениями рН, являются бедными по содержанию биофильных элементов), либо численность актиномицетов в почве сопоставима с количеством этих организмов в лесной подстилке.
В минеральных горизонтах обследованных лесных почв вниз по профилю численность актиномицетов постепенно уменьшается. Это особенно заметно при анализе профильного распределения актиномицетов по их минимальным значениям. В экологическом смысле нижний предел численности отражает пул микроорганизмов в почве. Величина пула не зависит от сезона, а определяется типом почвы. При сравнении различий биогеоце- нотических горизонтов по максимальным значениям численности мицелиальных прокариот наблюдается сглаживание различий между горизонтами почвенного профиля, и лишь нижние слои подстилки и верхний почвенный горизонт отличаются от остальных более высокой численностью актиномицетов.
Таким образом, горизонтами с максимальной концентрацией актиномицетов являются лесные подстилки и верхние почвенные горизонты (А1). Для малогумусированных бедных минеральными элементами песчаных подзолистых почв хвойных лесов подстилка является ярусом, максимально насыщенным актино- мицетами.
По вертикально-ярусной структуре лесных биогеоценозов изменяется не только численность, но и таксономический состав актиномицетов (рис. 118, 119).
Рис.
118. Распределение
актиномицетов по ярусам лесного
биогеоценоза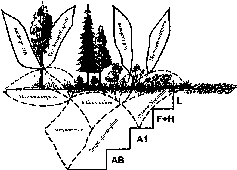
Рис.
119. Распределение
актиномицетов по горизонтам
дерново-подзолистой почвы
В филлосфере лесных растений обитают представители родов Streptomyces, Actinoplanes, Thermomonospora, Saccharopolyspora, Microbispora, Actinomadura, Micromonospora. Очевидно, актиномицеты нельзя считать постоянными обитателями филлосферы, что связано с экологической стратегией, присущей актиномицетам, которые при наличии в субстрате легкодоступных веществ не оказываются в первом эшелоне r-стратегов и для которых значительное количественное увеличение, а иногда и явное доминирование отдельных представителей, наблюдается обычно тогда, когда создаются условия для использования сравнительно труднодоступных (медленно разлагаемых) субстратов. С другой стороны, актиномицеты, являясь, как все прокариотные организмы, потомками древних обитателей Земли, несомненно, имели возможность вступать в разнообразные взаимоотношения с высшими формами жизни, в первую очередь с растениями на всех стадиях эволюции последних. Многие представители группы ак- тиноплан способны усваивать растительные пентозаны в качестве единственных источников углерода.
Доминантами среди актиномицетов во всех ярусах лесных биомов являются стрептомицеты. Исключение составляют слои моховой подстилки ельников и нижние горизонты (F и Н) подстилки в лесных биогеоценозах, где доля микромоноспор в актиномицетном комплексе сопоставима с долей стрептомице- тов. Представители родов Streptoverticillium, Micromonospora, Nocardia обнаруживаются в подстилке чаше, чем в минеральных горизонтах почвы. В лесной подстилке обнаружены представители рода Actinomadura.
Низкие значения рН лесных подзолистых почв способствуют развитию в них актиномицетов рода Streptosporangium, многие представители которых выдерживают кислые условия среды.
Использование показателей экологического разнообразия — частоты встречаемости и доминирования — выявило, что стрепто- мицеты встречаются повсеместно во всех ярусах лесных биогеоценозов, микромоноспоры — часто во мхах и нижних слоях лесной подстилки, где они могут доминировать; стрептоспорангиальные актиномицеты выявляются в основном в почвенных горизонтах, что, очевидно, связано с их фенотипическими признаками, в частности, способностью сохраняться в условиях низких значений рН среды.
Видовой спектр доминирующих в лесных биомах стрептомице- тов наиболее разнообразен в нижних слоях лесной подстилки. Здесь доминируют стрептомицетные виды, относящиеся к секции Cinereus серии Achromogenes; секции Albus серий Albus и Albocoloratus; секции Helvoloflavus серии Helvolus; секции Imperfectus.
Виды почвенных актиномицетов лесных биогеоценозов функционируют в условиях, где основными средообразующими факторами являются низкие значения рН среды и невысокое содержание органических веществ. Эти условия определяют заполнение основной части пространства экологических возможностей (65%) в почве леса только стрептомицетами секции Cinereus серии Achromogenes. Для почвы характерны стрептомицеты видов, принадлежащие к секции Cinereus серии Chrysomalius, секции и серии Albus Albus, секции Imperfectus.
В лесной подстилке чаще обнаруживаются представители Micromonospora fulvopurpurea, а в почве — Micromonospora aurantiaca и М. fulvopurpurea.
Результаты исследования актиномицетных комплексов лесных экосистем позволяют сделать заключение о специфичности комплексов, отличающей их от комплексов мицелиальных прокариот других ландшафтов гумидной и аридной зон (рис. 120).