

3.1. Гормон зависимая система транспорта ионов кальция в клетках.
При связывании определенных гормонов с рецепторами на поверхности клетки активируется путь регуляции поступления в клетку ионов Са2+ посредством инозитол 1,4,5-трифосфата (IP3). Данные о потоках кальция в компартментах клетки, которые участвуют в формировании IP3 - зависимых процессов, приведены на рис. 8 (см. на стр 11 данного текста).
Исследователи часто используют Ca2+-чувствительные флуоресцентные индикаторы для наблюдения за цитоплазматическим Ca2+ после IP3 - активации. Всплески Ca2+ отражают локальное открывание в эндоплазматическом ретикулуме (ЭР) отдельных Ca2+-каналов или их небольших групп, когда IP3 связывается с рецептором на мембране ЭР.
Поскольку различные Ca2+-связывающие белки служат буфером Ca2+ и ограничивают диффузию Ca2+, сигнал часто остается локализованным в месте входа Ca2+ в цитозоль. Однако, если внеклеточный сигнал достаточно силен и продолжителен, этот локальный сигнал Ca2+ может распространяться по цитозолю как самоподдерживающаяся волна Ca2+, во многом напоминающая потенциал действия в аксоне. За такими всплесками концентрации Ca2+ часто следуют следующие всплески, длящиеся обычно несколько секунд. Колебания Ca2+ могут продолжаться столько времени, сколько рецепторы на поверхности клетки остаются активированными. Считается, что, по крайней мере, частично, волны и колебания зависят от сочетания положительных и отрицательных обратных связей между кальцием и IP3-рецепторами и рианодиновыми рецепторами: высвобождаемый Ca2+ сначала стимулирует дальнейшее выделение Ca2+ обоими рецепторами в процессе, носящем название Ca2+-индуцированного высвобождения кальция; но затем, как только его концентрация становится высокой, Ca2+ ингибирует высвобождение, и такая отрицательная обратная связь с задержкой приводит к колебаниям.
Наша цель в данной задаче – изучить на модели возможность устойчивых колебаний в системе потов ионов кальция.
3.2. Модель гормональной регуляции потоков ионов кальция в клетке.
Минимальная модель потоков ионов Ca2+ в клетке оперирует двумя переменными S1 для кальция в цитозоле и S2 в гормон нечувствительном депо (гормон-не-чувствительный пул). На схеме рис. 8 концентрациям S1 и S2 соответствуют обозначения Ca2+Cyt и Ca2+депо.
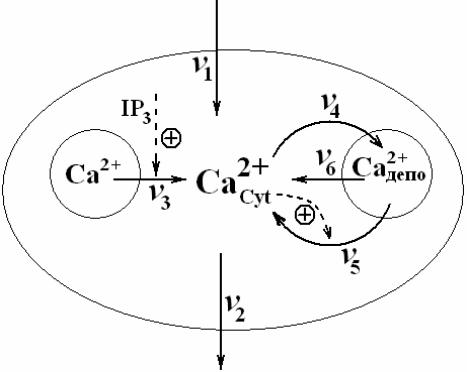
Рис. 9. Схема инозитол (IP3) зависимых потоков ионов Ca2+ в клетке. Обобщение данных, показанных на рис. 7, для формирования минимальной модели потоков ионов кальция. Обозначения метаболитов и скоростей см. в тексте.
Модель включает инозитол (IP3) зависимые потоки ионов Ca2+ на границах компартментов, показанных выше на рис. 8: внеклеточная среда, цитозоль клетки, гормон-чувствительный ЭР и депо гормон-не-чувствительного ЭР. В модели сопоставим этим потокам скорости, формирующие баланс метаболитов S1 и S2 .
Приток и отток цитозольного кальция через плазматическую мембрану со скоростями v1 и v2.
Гормонально активируемое высвобождение кальция из внутриклеточного пула, чувствительного к горомону (инозитол-три-фосфат IP3) (v3).
Активный транспорт цитозольного кальция в гормон-не-чувствительный пул (v4).
Высвобождение кальция из гормон-нечувсвительного пула, которое активируется цитоплазматическим кальцием (механизм Ca2+-индуцированного высвобождения кальция, скорость v5).
свободный отток кальция из гормон-не-чувствительного пула в цитозоль (v6).
Модель состоит из двух дифференциальных уравнений:
![]() (8)
(8)
Внеклеточная концентрация Ca2+ и значительно превышает цитозольную и полагается постоянной (буфер). Поэтому скорость потока v1 неизменна. Более того, подробное рассмотрение системы (8) [ссылка] позволяет скорость v3 считать постоянной и объединить эти две скорости в модели v0 =v1 + v3. Скорость v3 постоянна по причине быстрого заполнения кальцием гормон-чувствительного депо ЭР. К тому же эта скорость зависит только от величины IP3 стимула в системе. Поэтому далее в исследовании модели скорость v0 можно принимать за параметр (интенсивность стимула), определяемый условиями эксперимента по стимулированию клетки.
![]() (9)
(9)
Рассмотрим детальные механизмы всех скоростей модели (9), используя для них достаточно простые предположения, как показано в таблице 1.
Таблица 1. Механизмы скоростей, предлагаемые для схемы процессов на рис. 8, 9.
Скорость |
Механизм |
Процессы, от которых зависит vi |
v0 |
v1 + v3 |
Утечка по градиенту концентраций на мембранах: плазматической (v1) и гормон-чувствительного ЭР депо (v3). Зависит от интенсивности стимула. |
v2 |
k2S1 |
Ca-АТФаза за счет гидролиза АТФ выкачивает ионы против градиента концентрации из клетки во внешнюю среду (v2) или в гормон-не чувствительное ЭР депо (v4) |
v4 |
k4S1 |
|
v5 |
|
Уравнение Хилла.
|
v6 |
k6S2 |
Утечка по градиенту концентраций на мембране гормон-не-чувствительного ЭР депо |
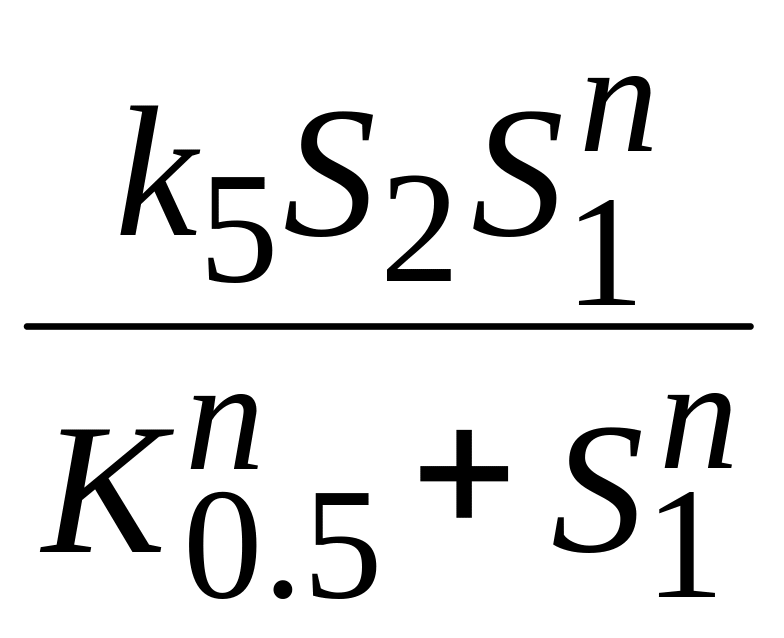
После подстановки конкретных описаний скоростей vi получим систему (9) для математической модели внутриклеточных колебаний концентраций кальция.
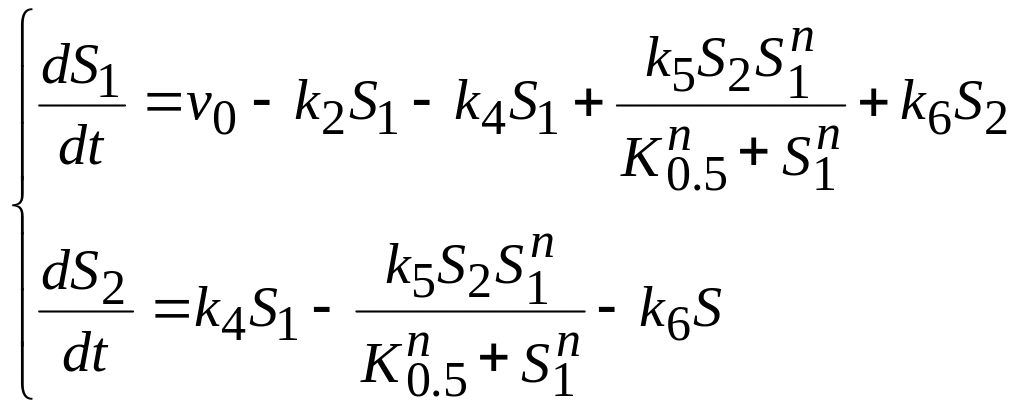 (10)
(10)
С помощью модели
(10)
можно исследовать поведение системы
потоков кальция в зависимости от величины
параметра
![]() (суммарного поступление кальция через
плазматическую мембрану и гормонально
активируемого высвобождения кальция
из внутриклеточного пула). Окончательно
суждение о существовании устойчивых
колебаний подтверждается численным
интегрированием системы уравнений при
условии выбора параметров из диапазона
неустойчивости.
(суммарного поступление кальция через
плазматическую мембрану и гормонально
активируемого высвобождения кальция
из внутриклеточного пула). Окончательно
суждение о существовании устойчивых
колебаний подтверждается численным
интегрированием системы уравнений при
условии выбора параметров из диапазона
неустойчивости.
Далее выполним задания для подготовки дома и для работы в классе на компьютере с целью исследовать поведение модели (9) в зависимости от величины параметра (суммарного поступление кальция через плазматическую мембрану и гормонально активируемого высвобождения кальция из внутриклеточного пула).
Дополнительное задание. Исследуем
систему двух дифференциальных уравнений
(8) в общем виде, не детализируя зависимости
скоростей vi
от концентраций S1
и S2.
Указание В стационарном состоянии
 и
и
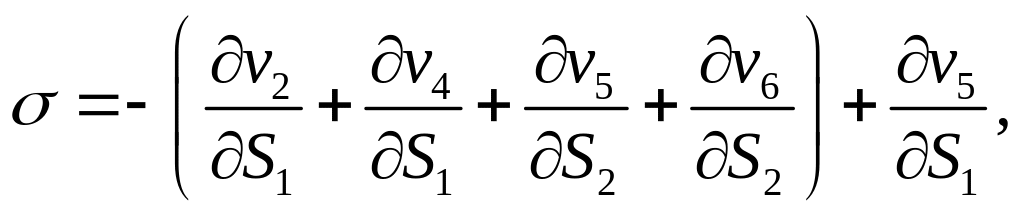
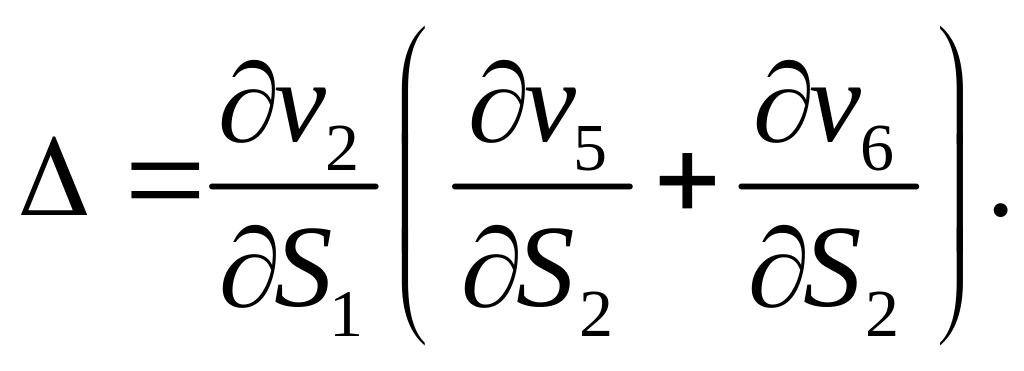 Вспомните исследование, проведенное в
разделе 1.3.1. и ответьте на Вопрос:
«Как должны зависеть скорости системы
(9) от переменных модели для того, чтобы
в системе могли существовать устойчивые
колебания?»
Вспомните исследование, проведенное в
разделе 1.3.1. и ответьте на Вопрос:
«Как должны зависеть скорости системы
(9) от переменных модели для того, чтобы
в системе могли существовать устойчивые
колебания?»
Разумно считать, что скорости v2 и v4 двух Ca2+/АТФ-зависимых насосов активируются цитозольным кальцием (т.е. зависят от концентрации S1), в то время как они не зависят от концентрации запасенного кальция S2. С математической точки зрения это означает, что:

(11)
Для скорости утечки v6 можно считать, что
![]() (12)
(12)
Вполне возможно, что это процесс обратимый, однако формально свободное поступление кальция в гормон-не-чувствительный пул можно учесть в потоке v4.
Исследуем систему двух дифференциальных уравнений в общем виде, не детализируя зависимости скоростей vi от концентраций S1 и S2.
В стационарном состоянии
Тип особой точки определяем исходя из коэффициентов линеаризованной системы:


Таким образом,
Поскольку нас интересуют ситуация,
когда в системе имеют место автоколебания
(т.е. существует предельный цикл), выпишем
необходимые условия
![]() и
и
![]() меняет знак. Поскольку в наших
предположениях
меняет знак. Поскольку в наших
предположениях
![]() ,
то необходимым условием бифуркации
Хопфа является неравенство:
,
то необходимым условием бифуркации
Хопфа является неравенство:
![]() Далее, для того, чтобы
меняла знак, необходимо выполнение
неравенства
Далее, для того, чтобы
меняла знак, необходимо выполнение
неравенства
![]() .
Это соотношение имеет чрезвычайно
важный смысл: оно означает, что в
рассматриваемой схеме активация
высвобождения кальция из
гормон-не-чувствительного пула цитозольным
кальцием (наличие положительной обратной
связи) — это необходимое условие
существования автоколебаний концентраций
кальция.
.
Это соотношение имеет чрезвычайно
важный смысл: оно означает, что в
рассматриваемой схеме активация
высвобождения кальция из
гормон-не-чувствительного пула цитозольным
кальцием (наличие положительной обратной
связи) — это необходимое условие
существования автоколебаний концентраций
кальция.
Рис. 8 Схема. Потоки кальция в компартментах клетки.

-