

54. История открытия и пространственная организация в клетках растений процессов с4-цикла.
Так, советский ученый Ю. Карпилов и австралийские ученые М. Хетч и К. Слэк выявили, что у некоторых растений, главным образом тропических и субтропических, таких как кукуруза, сахарный тростник, сорго и другие, основная часть меченного углерода (14СО2) уже после нескольких секунд фотосинтеза обнаруживается не в фосфоглицериновой кислоте, а в щавелево-уксусной (ЩУК), яблочной (ЯК) и аспарагиновой (АК) кислотах.
В этих кислотах можно обнаружить в первые секунды до 90 % поглощенного 14СО2. Через 5–10 минут метка появлялась в фосфоглицериновой кислоте, а затем в фосфоглицериновых сахарах.
Так как эти органические кислоты содержат 4 атома углерода, то такие растения начали называть С4-растениями в отличие от С3-растений, в которых радиоуглеродная метка появляется прежде всего в ФГК.
Это открытие положило начало серии исследований, в результате которых подробно был изучен химизм превращения углерода в фотосинтезе в С4-растениях.
Еще более интересны отклонения от типичного строения листа, влияющего на фотосинтез, связаны с генетическими различиями. В таких растениях, как кукуруза, сахарный тростник (С4-растения), каждый проводящий пучок окружен одним слоем крупных клеток хлорофиллоносной паренхимы, образующих обкладку проводящего пучка. Фотосинтез идет и в клетках мезофилла и в клетках обкладки проводящего пучка (рис. 2.2).
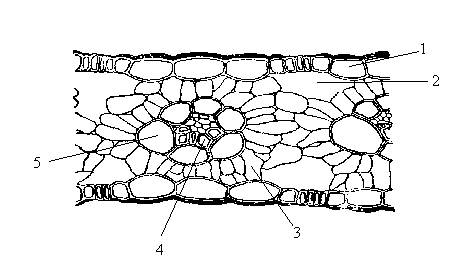
Рис.2.2. Структурные особенности С4-растений: поперечный разрез листа кукурузы:
1 – эпидермис; 2 – межклетники; 3 – мезофилл; 4 – проводящий пучок; 5 – клетки обкладки
Морфология клеток листа С4-растений:
– многочисленные воздушные полости, по которым воздух из атмосферы подходит ближе к большему числу фотосинтезирующих клеток, обеспечивая более эффективное поглощение углекислоты;
– слой клеток обкладки сосудистых пучков, плотно упакованных около проводящих пучков;
– клетки мезофилла, которые располагаются менее плотными слоями около клеток обкладки сосудистых пучков;
– большое количество плазмодесм между клетками обкладки сосудистых пучков и клетками мезофилла;
– содержание как гранальных, так и агранальных хлоропластов, которые отличаются и по характеру проходящих в них реакций фотосинтеза.
55. Отличие малатного и аспартатного путей у с4 – растений. Схема реакции малатного пути.
В настоящее время в зависимости от того, какая из органических кислот (малат или аспартат) декарбоксилируется, С4-растения делят на два типа: малатный тип (кукуруза, тростник) и аспартатный (сорго и др.) типы.
В свою очередь растения последнего типа подразделяют на две группы: растения, которые используют в реакции НАД-зависимую малатдегидрогеназу и растения, которые используют фосфоенолпируват-карбоксикиназу.
Как отмечалось, С4-растения отличаются от С3-растений и по анатомии листовой пластины. Фотосинтез идет в клетках обкладки и в клетках мезофилла. Оба типа фотосинтезирующих тканей отличаются по строению хлоропластов. Хлоропласты клеток мезофилла имеют строение, присущее большинству растений: они содержат два типа тилакоидов – тилакоиды гран и тилакоиды стромы (гранальные хлоропласты). Клетки обкладки содержат более крупные хлоропласты, часто наполненные крахмальными зернами и не имеющими гран, т. е. эти хлоропласты содержат только тилакоиды стромы (агранальные).
Считают, что агранальные хлоропласты образуются в процессе онтогенеза листа из обычных гранальных хлоропластов, так как на ранних стадиях развития эти хлоропласты также имеют граны.
Таким образом, для С4-растений характерны следующие особенности в строении:
– многочисленные воздушные полости, по которым воздух из атмосферы подходит непосредственно к большому количеству фотосинтезирующих клеток, обеспечивая эффективное поглощение углекислоты;
– слой клеток обкладки сосудистых пучков, плотно упакованных около проводящих пучков;
– клетки мезофилла, которые располагаются менее плотными слоями около клеток обкладки сосудистых пучков;
– большое количество плазмодесм между клетками обкладки сосудистых пучков и клетками мезофилла;
– содержание как гранальных, так и агранальных хлоропластов.
Для малатного типа растений характерен следующий путь фотосинтеза. Гранальные и агранальные хлоропласты отличаются и по характеру проходящих в них реакций фотосинтеза. В клетках мезофилла с мелкими гранальными хлоропластами происходит карбоксилирование ФЕП с образованием ЩУК (первичное карбоксилирование), а затем образуется малат. Малат перемещается в клетки обкладки. Тут малат окисляется и декарбоксилируется с участием малатдегидрогеназы. Образуется СО2 и пируват. СО2 используется для карбоксилирования рибулозо-1,5-дифосфата (вторичное карбоксилирование) и таким образом включается С3-цикл, который идет в агранальных хлоропластах клеток обкладки. Пируват возвращается в клетки мезофилла, где фосфорилируется за счет АТФ, что приводит к регенерации ФЕП, и цикл замыкается
56. САМ-цикл, его роль в приспособлении к засушливым условиям существования. Сравнение САМ-цикла и С4-цикла.
Кислотный метаболизм толстянковых (КМТ или САМ цикл).
для САМ-растений характерны следующие особенности:
– устьица обычно открыты ночью (в темноте) и закрыты на протяжении дня. Такое состояние устьиц противоположно тому, что характерно для других растений;
– фиксация СО2 происходит в темный час суток в содержащих хлоропласты клетках фотосинтезирующих тканей листа или стебля, причем синтезируется значительное количество свободной ЯК;
– ЯК кислота накапливается в больших вакуолях, которые характерны для клеток этих растений;
– на свету ЯК декарбоксилируется и освободившийся СО2 превращается за счет энергии света в сахарозу и запасной глюкан (например, крахмал) в световых реакциях фотосинтеза С3-типа.
– в последующий темновой период часть запасенного глюкана разрушается с образованием молекул-акцепторов для темновой реакции фиксации СО2.
Следовательно, в тканях САМ-растений существует суточный цикл: ночью содержание запасенного глюкана падает и содержание яблочной кислоты повышается, а днем происходят противоположные изменения. Следовательно, реакции разделены во времени, а у растений С4-типа процессы разделены в пространстве.
С4 и САМ-типы фиксацин СО2 являются дополнительными, облегчающими функционирование цикла Кальвина в растениях, которые приспособились к жизни в условиях повышенных температур (сахарный тростник, сорго, кукуруза и др.), или недостатка влаги (суккуленты).
В заключении можно дать следующую характеристику процесса фотосинтеза:
– сущность фотосинтеза состоит в фотоокислении воды с выделением кислорода и использованием водорода на восстановление СО2 с образованием органических веществ;
– фотосинтез состоит из двух фаз: световой и темновой;
– энергия поглощенного света используется для движения электронов против градиента окислительно-восстановительного потенциала;
– транспорт электронов сопряжен с синтезом АТФ;
– световая стадия происходит в тилакоидах стромы и гран, а темновая – в строме хлоропластов, т. е. для световой стадии необходимы мембраны.